* 生物棟停電(7/4fr18:00〜7/6sn16:00)
90 モデルの実装(6月中完了を目処に)
90 trade-offについての検討(7/6迄)
80 モデルの空間拡張(7月中旬完了を目処に)
30 ECOLOGICAL MODELLING 3rd ed. 適当にサマライズ。
10 Benard流を組み込んだモデルの設計と評価
10 sink→resourceモデルについて調べ物と検討。
? プリントサーバの構築
午前中読書ののち私用、予約特典に惹かれて予約してきました…。午後読書したり調べ物をしたりモデルについて考えたり。
Amazonより、先日注文した『Ecological Niches』届く。
某氏が中身をぱらぱらと眺めて「thoreticalのフレーズをタイトルのどこかに入れるべきではないのか。」などと宣う。然り、これはそういう本なのです。
『個体群生態学』『Fundamentals of ECOLOGICAL MODELING』と併せて、読んでない本が増えて行く。消化しないとね。
夜半過ぎより某氏の誕生祝(の名目で)研究室で黒糖焼酎をいただく。奄美の御土産で旨い酒でした。
ミジンコの一般的な体サイズについて聞いたのでメモ。
μg・Cで、adult / egg(or juvenile) = 10/1〜10/3、adult = 10〜30くらいを大まかな目安にしておくと、大体そんなものらしい。この種の大まかな数値の目安をちょいちょいと聞けて即答で返ってくるという状況は実にありがたいものだ。
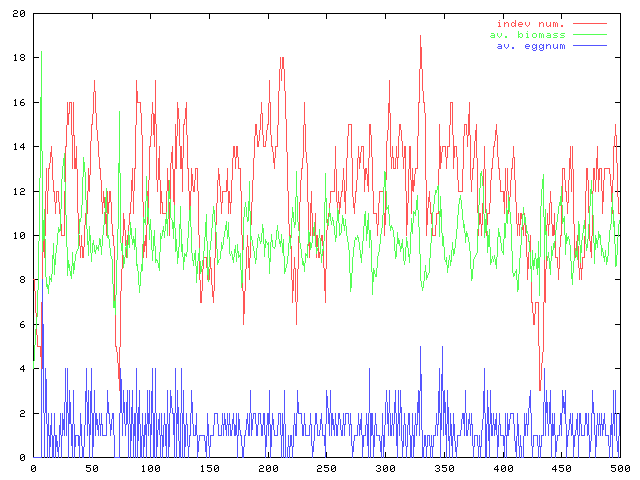
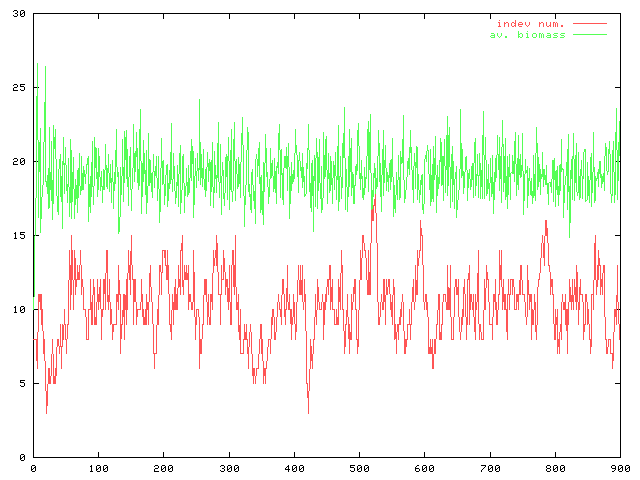
んで、adultのサイズを20に固定して卵サイズのadultに対する相対的な大きさの違いが個体数の変動と平均バイオマスにどのような影響を与えるかについてのグラフ(赤:個体数、緑平均バイオマス)。…よく見ると両方のグラフで縦軸の縮尺が違っている。こういうのはミスリードを招きやすいので注意しておかないと。
卵サイズが大きいとき(egg=4)に比べると、卵サイズの小さな条件(egg=2)のほうが個体数変動の振動が大きい。これは繁殖一回当たりの再生産量が大きいために、その分変動幅が大きくなっているからだと考えられる。
1.個体数の母集団を大きくする、2.死亡率を下げる、の二つによって個体数の振動は(相対的には)比較的小さくなるようだ。ま、それはそうだわな。
モデルでの個体の死亡の扱いの続き。
昨日の問題をもう少し考えてみるべく、教官殿に個体の間引き方について聞いてみたところ、さらりと答えてくれたのだった。結局のところ個体ベースモデルにおいては
で、これからやること。
取り急ぎ、間引きに関してはモデルのデザインを考えるとして、平行して藻類の形質セットに関するtrade-offについての資料集めと整理と統計解析。trade-offがあるかないかは別として、これをまずはじめにやらないと計算の始め様が無い。
同時に個体ベースモデルの格子モデルへの拡張も進める。前者を今週中を目処に仕上げ、後者については空間無しについて計算させながら、ぼちぼちとまあ7月半ばくらいを目処に。
add TODO stack:
+ trade-offについての検討(7/6迄)
+ モデルの空間拡張(7月中旬完了を目処に)
梅雨の合間、束の間の良い天気であった。午後より秋保大滝付近を軽く流して、夕刻よりコーディングの続きとか。
先日の個体の死亡の扱いについて続き。
この前の式はちょっと考えればすぐにわかるのだが、寿命に達するまで絶対に死なないのだった。したがって没。
ではどうするかということで、齢構成分布に従って一様な死亡率と繁殖率を与えてみる。このときどうなるかというと、個体数(N)と成長率(g)と死亡率(m)をもちいてdN/dt=(g-m)N、したがってN=N0exp{(g-m)t}となる。本当はこれに資源に対するfunctional responseが導入されて飽和(g>rで無限に増加しない)するわけだが、ここでは省略。
で、これを実際に個体ベースモデルの中で表現するためには、結局ランダムサンプリングで間引くしかないような気がする。母集団を増やすことで解決されると期待して次に進めて…良いのだろうか。
一日中自宅で読書など。雨もひどいので良しとする。
午前中お仕事、午後時間を置いてバイト、夕刻より輪読会。その後現状で出てる結果を見せる。
モデル作業。
現時点での結果を見せたところ、ミジンコの個体にかける死亡の効果をランダムにかけている点について、効果が散らばってしまうからよくないという話になる。
つまり、移動の段階でランダムな移動の効果を見たいためにはここでランダムに死亡するような形はよろしくなかろう、ということ。で、齢構成に従うとか何とかいろいろ考えた果てに死亡率が時間とともに増加し、ある値を超えたところで決定論的に死亡するという形にすることに。
どのようなデザインで設計するかが問題になるが、どうしようかね。
夕刻まで輪読会(向こう)の準備。夕刻こちらの輪読会。夜コーディング。
逐次訳は効率が悪く、意外と時間がかかる。流し読みなら30分とは掛からない程度の内容なのに。
陸水学会の申し込み取りやめ。
向こうの先生のアドバイスによると、この種の研究は陸水学会では「ふーん」というので終わりがちで発表事態が目的でなければ(研究上で何らかのアイデア等を求めるなら)あまりメリットはないだろう、とのこと。
それはあまり面白くないねえ、ということで陸水学会は見送り。目先の目標がなくて気が抜けました、なんてことのないように引き締めていこう。
コーディングの続き。
格子モデルの単位となるユニットが完成(一応)。まずはそれを使って空間を考えない条件での計算を行うようなプログラムの作成。進んでいるのだが、手の遅さは如何ともしがたい。
del TODO stack:
− 陸水学会申し込み
午前中セミナー、午後勉強会。
コメンテータとしての機能が不完全で、将来的な課題をたくさん残す結果になった。
コーディングのほうはがりがりと。ミジンコ部分はあらかた完成。格子の単位についてガリガリ…
午前中自宅にて読書。友人から借りた「歌の翼に」(菅浩江)読了。
相変わらず心の闇を描かせたらこの人は巧い。後半かなりテーマ的にも重たい話なのだが、ドロドロとしたものをあまり感じなく読めるのはこの人ならではと思う。久々によい本を読ませてもらった。よい本を読むと何より気力が充実する。テーマがよく似ているという「永遠の森」も文庫か新書にされないものか。
Amazonで調べたところ「プレシャス・ライアー」という新刊が出ているようで、カートに登録だけしておく。
新しいノート所感
新マシンはMP3聞いてPDFを見ながら適宜WEBで調べ物しつつMS-Wordで文書作成なんてことをやってもぜんぜん平気なのだった。うむ、快適。今まで使っていたノートは後輩の学生氏に15kにて譲る。それにしても5千円で買い叩けというのは幾らなんでも足元見すぎです。>教官殿。
相変わらずがりがりとコーディングの続き。手が遅い。
すっきりとまとまらないので基底クラスにも少しだけ手を入れることに。ああ、OOPを逆行するようなコーディングスタイル…。金曜日に結果を持って行きますなんて口約束してしまったので頑張らないと。
IBM化は一応の形が出来上がったので、統括するフレーム部分を書いてβ版、かな。フレーム部分はこれがそのまま格子モデルのセルとしても機能するようにするので、作業量は結構ある。まあ以前書いたものがかなり流用できるんではなかろうか。それが終わり次第、条件を用意して計算開始。
論文よみ
動物プランクトンの個体ベース化について論文が出ている。Zadereev E.S, et al.(2003), Ecol. Model. (162), pp15-32。うーむ、Ecol. Model.あたりに投稿する研究者なら普通に思いつくアイデアということか。作業中にちょっと出鼻をくじかれたような気分であるが、まあ仕方ない。研究テーマが違うんだからよしということにしよう。
後できちんと読んでおくことにする。
陸水学会の申し込みについて。
教官殿にどないでしょーと振ってみたところ、向こうの先生も交えて研究計画の話をしようという話から、はよ結果出して進捗状況よこせ、と。前回出した分は読まないつもりかな。申込締切から逆算して、後6日以内には取り敢えずの結果を出さないと二進も三進もいかなくなるようだ。むー。
水曜日のセミナーで司会兼コメンテータを任される。
それは有難いのだが、内容がやや特殊でどう突っ込んでいいのかわからない。そもそもコメンテータって何するんですかという情けない有様。発表者から進め方についての指針をメールでもらって何とかする。
やることが多いのでどれかひとつに専念できないという状況が長引く。これからはずっとそうだろうから慣れないとね。
研究室で使っているノートPCが新調(先輩のお古ですが)
これでまた10万の借金生活。いろいろと環境を整えるのに午後いっぱい掛かる。今日はこれでおしまいということにする。
午後起床。コーディングの続きとか資料の整理とか。
紹介された本
"Competition and Coexistence", Sommer U.(2002), Springer(2002), Ecological Studies(161) をAmazonにて買おうとするも1万円超えて断念。実にいい本なのに高すぎ。必要な個所だけ複写させていただくことにする。
もう一冊、"Eclogical Niches: --", Chase J.M., and Leibold M.A. の方は購入。Amazonにてギフト使用で2500円、Paperbackなら安い。
午前中仕事、午後仕事。夕刻より輪読回。逐次訳だと思いしっかりと下準備をしなかったのは失敗だった。向こうの学生さんに迷惑を掛けてしまい申し訳なく思う。つぎから説明に詰まる個所についてはきちんと対処すること。
向こうの研究室に顔を出して話をすると否応無しに危機感が募るので、発破をかける意味合いでは良いのだ、が、心理的な焦燥感は結構在る。という分析できるくらい落ち着いているのか。先を急ごう。
といいつつ修士過程のヒトの「キーボードの配列がおかしいんですけど。」という質問に対処したりとか。なんでus設定に日本語106挿して使ってるかな。かれこれ3,4ヶ月はそのままの設定のはずだったはずだけど。いわく、「今まで使ってなかったから」。なはは。
先日の玩具を使った思考実験(?)の続き。
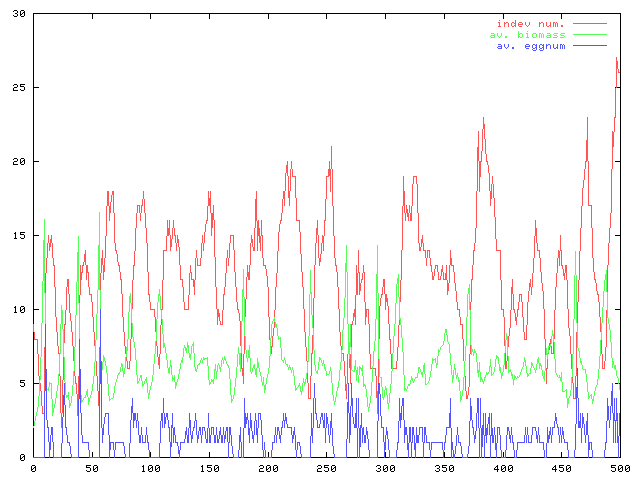
逐次卵が出来次第個体が増えるのはミジンコ的におかしいという指摘を受け、一定期間卵を抱える(=ある時点で数個の卵を生産)という形に変更。加えて卵の生産戦略が違うときの動態について、個体数(赤)と平均のバイオマス(緑)と平均の卵生産数(青)の時間変化。卵生産に満たない余分な資源を次回の生産に持ち越すときと親の資源を使ってもうひとつ卵を作るときのグラフ。前者ではバイオマスが高めで個体数と卵数が少なめ、後者ではバイオマスはバイオマスは低いが個体数と卵数は多目という至極ありきたりな結果である。
現状で体サイズ(=バイオマス)が大きいと生存率が上がるとか考えない限り、利用資源量が同じなら(個体数×バイオマス=一定)ということになるだろう。つまり、そこにあれこれ条件をかけないのであればどちらの戦略でも結果は変わらない、ということか。
ただし問題になるであろう点がひとつ。排泄によるリサイクル量はバイオマスの関数になっているので、卵生産戦略の違いによってリサイクルの効果の大小が変わってくるような気がする。
昼過ぎ起床。新パッチがでたゲームを明け方まで遊んでいたのが理由なんだが、平日はいかん。反省。
午後、明日の輪読回の準備や、久しぶりにオーストラリアから帰ってきた同期と話したり。英語が堪能になっているのと、研究と将来についてのモチベーションには僕も頑張らねばという気にさせられる。
夕刻より輪読回。後半の演者のユーモアあふれる進行のおかげで久しぶりに面白い輪読回であった。
昨日(18日)より、陸水学会の講演申し込み開始。受付は7月3日14:00までなのでそれまでに済ませないと。
発表の申し込みや要旨の提出などは、生態学会のときはいまどき所定の紙に書いて郵送もなかろうと思ったのだが、それと較べると徹底していて且つ洗練されていると思う。
要旨原稿の指定を見るに、つまるところ8月の頭くらいまでには結果と考察を出さないとまずいわけね。頑張ります。
午前中研究室のセミナー、午後セミナー、夕方まで勉強会。(おいおいおい…)
疲労困憊を言い訳にして今日はコーディングしないで読書。
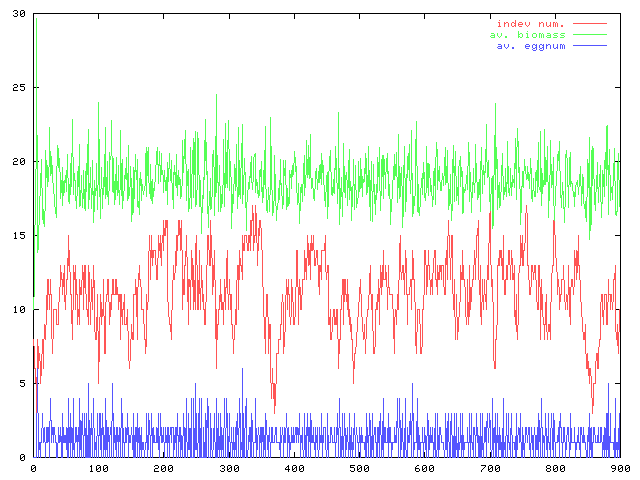
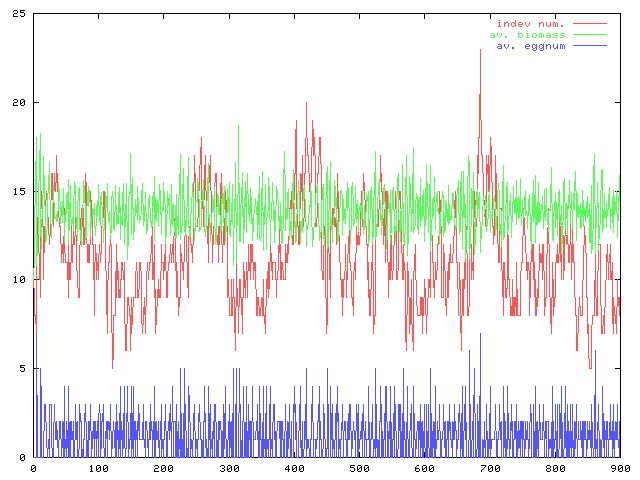
コーディングの続き。モデルの卵生産の部分は雛型ができた。個体数(赤)と平均のバイオマス(緑)の時間変化と資源量の時間変化のグラフ。パラメータは適当だが形としてはこんなもんだろう。
GCC-STLのコンテナに使うerase()について初歩的な勘違いしていたことが判明。下記のように書いて失敗。
vector::iterator i;
// 値が3なら削除
for (i = x.begin(); i != x.end(); i++)
if (*i == 3) x.erase(i);
vector::iterator i;
// 値が3なら削除
for (i = x.begin(); i != x.end(); ){ // ここではiに何もしない
if (*i == 3) x.erase(i);
else i++;
}
本を読んだり調べものをしたり雑用を片付けたりする。そういうのは後でいいからさっさと手を動かすべきなのは分かっているのだが、他にやることがなくなるか追い詰められるまで作業に掛からない性分らしい。
とはいえ本格的にそれ以外の作業が片付きつつあるので実装開始。内的・外的要因の両方により、邪魔されずにペースを維持するのが一番大変だったりする。つまるところ集中力が足りない、ということなのだが。
教官殿との作戦会議は延期。例によってまだ読んでないとの返事であった。どのみち現状の説明がメイン(と思う)ので、放っといて作業を進める。
動物プランクトンIBMの雛型をコーディング中。作業に入ってしまえばいろいろ問題は出てきてもそれはそれで楽しい。シミュレーションモデルには書いてみないと固まらない部分というのもあるわけで、このペースで明日以降も続行続行。
休日ということにして、プリントサーバ用のマシンのセットアップをしてました。smbとnetatalkを入れたところまで。共有設定とかはケーブルを買ってきてプリンタを繋いだ後でします。
正午起床。文献複写のため図書館へ行くも目当ての雑誌(Annu. Rev. Ecol. Syst.)は置いてなく、実は電子ジャーナルで利用できた。
いつものことながら、モデルを実装する作業は始める前に色々と手続き(というか儀式)をしないとどうもやる気が揚がらない。で、kernelのいれかえやら始めてしまうのだった。
午前中仕事、午後私用と雑用をはさんで仕事。火曜に修理に出した計算機がもう帰ってきたのだった。電源部のみ交換とはいえすばらしー速さですよー。年内保証で費用の発生も無し。
夕刻より輪読回。毎回一人ずつの分担を決めて進める形式のようで、次回からは僕も担当。とはいえたいした量ではない(4p,net)のでさして時間は取られないだろう。
昨日のtrade-offについて。
Coijman(2000)からヒントを得て、良いアイデアを思いつく。窒素とリンの両方の基質が飽和濃度に達しているときの最大吸収速度をv1,v2、対応する膜上のチャンネルの個数をn1,n2、透過性(親和性)をa1,a2とし、
v1=a1*n1n1+n2=C(一定)と仮定して、v1とv2について整理すると、
v2=a2*n2
v1=(a1/a2)*(C'-v2) [C'は定数]ここでa1,a2は膜上のチャンネルの透過性なので生物種間で違いはないため、a1/a2は定数(R比よりK>1)である。よって、藻類種iのv1とv2は以下の関係式で与えられ、線形のtrade-offとして表される。
vi1=K*(C'-vi2) [C'は定数]vi1とvi2にトレードオフがあるならば、同じ分類群の藻類ではKとC'はほぼ同じ値になっているはずである。過去の論文のvi1とvi2についてのデータからKとC'の実際に近い値を求めることができる。窒素とリン:Andersen(1997),Springer(1997)や窒素と珪素:Tilman, Kilham and Kilham(1982), Annu. Rev. Ecol. Syst. (13), pp349-372の他、いろいろな文献が集められるだろう。
打ち合わせのために進捗状況とモデルの詳細な構成をメールで教官殿へ。月曜日に打ち合わせ。
聞くところによると、どうやら研究に関して教官殿に心配をされている様子。そろそろ何とか名誉挽回を図りたいものだ。
午前中自宅で読書、消化量が増分に追いつかない…。午後雑用、論文読んだりトレードオフの関数形を実際に書いてチェックしたり。夕刻輪読回。
研究室内で使われているIPとethアドレスの管理表の作成。
zshとperlのおかげで作業自体は一瞬(返事が返ってきたアドレスのarpを取るだけ)なのだが、電源の入ってないマシンには無力…。電源を入れて回って一括でデータ回収すれば良いのだが、結局誰がどれを使ってるかを調べなければいけないのでethアドレスを手で入力しないですむ程度にしか手間は減らないのだった。
教官殿の部屋以外はほぼ回収完了したが、実はそこが最難関だったりする。
夕刻、教官殿より月曜に研究計画の打ち合わせをする旨、口頭にて通達。
予定通り明日中に宿題(前回の問題点への回答)含めた資料を用意する。ほぼ完了しているので速度的な問題は無し。いま取り組んでいるモデルの論文用introductionとmethodのアウトラインを、来月中に提出することを視野に入れると、提出資料の内容はまだ何箇所か詰めが甘いように思う。が、そのあたりは指摘されるのを待ってみる。
Muller et al. (2001), Ecol. Letters(2001), 4, pp519-529を読む。
生産に掛かる不足資源の律速をLiebig's Minimumをとる従来の不連続(discontinuous)な関数から、調和平均likeな連続(continuous)の関数に替えた、という部分が主旨。
Liebigの方だと独立の2変数の最小の方を選択するため、その間のtrade-offは別に記述せざるを得ない。これに対して、2種の資源の律速効果をひとつの連続式で記述すると、ある生産速度下における資源の取り込み速度のtrade-offを表現できる。当初は何が凄いのかわからなかったが、理解すると実にエレガントである。
2種の資源の取り込み速度のtrade-offについて。
生理的な側面から考えると、吸収速度について細胞膜表面上の結合部位の数を有限と考える(これは細胞膜の表面積が有限と考えても同じ)。つまり資源の最大吸収速度の間にはtrade-offが生じると考えられるわけだ。
しかし問題は、最大吸収速度の間にどのような形でtrade-offをかければよいのか分からない。もう少し考えたり相談したりしないといけない。
午前中セミナー、午後勉強会。
手を動かすためにはもう何本か論文や本を読む必要があることが判明。論文を読んだり資料を探したり、と。
午前中だらだらと雑用私用。午後資料の纏めとモデルについて打ち合わせ。
モデルについての話し合いの結果。恐ろしい勢いで話が進み、個体ベース案とトレードオフの両方について一先ずの見通しが立ってしまった。つまるところ作業は第二段階に移行。明日から(にする)は大忙しだ。
以下、話し合いの内容を忘れないうちに簡単にサマライズ。
まずは昨日のモデルにおける拡散の扱いについて。
昨日の案についてはあれで良いのだが補足として:
個体ベースの扱い方について。
拡散などに関しては問題らしい問題は無し。問題は個体の増え方に関してで、こちらが持っていったアメーバ増殖likeなアイデアはあまりreasonableでないということで、あっさり没。ではどうするかということで出たのが生産卵数を純生産/卵サイズで与える(b=NP/E)案。こちらは割合にすっきりとまとまっていてよい感じである。ただしこちらのアイデアでは、生活史戦略というか卵生産戦略自体が何某かの影響を持ってきそうな構成である。
どのような戦略が在りうるかということで適当にピックアップしてみると、
E=(一定)とするとき、
動物個体の移動速度と競争能力(生産性)のトレードオフについて。
当初、移動性×生産性=一定という関数形を考えていたのだが、移動性と生産性の間の関係の記述としては些かおかしいという指摘を貰う。で、移動に掛かるコストが生産性から引かれるという形で落ち着く。
競争者(PP)の栄養塩要求性についてのトレードオフの扱いについて。
栄養塩の吸収速度が細胞の表面積に比例し、Resource/Carbonの比率が小さく(=carbon rich)なると細胞の嵩が増すと考えると、吸収速度とcell quotaの間にトレードオフが生じると考えられる。さらに、吸収速度について細胞膜表面上のチャンネルの数をほぼ一定と考えると、N親和性とP親和性の間にトレードオフが生じる。この両者によって、吸収速度、cell quota、N/P要求性を纏めて扱えることになる。
この辺の生理的メカニズムからの説明とモデルにおけるトレードオフについては、Muller et al. (2001), Ecol. Letters(2001), 4, pp519-529 および、Kooijman(2000), Cambridge Univ. press(2000)に詳しい(ようだ)。読んでおかないとなあ…。
形質の開始時の組み合わせについて、ある形質から両方向に広がる新規ニッチ開拓型の組み合わせ(というか種数増加)と両極端な形質間を飛び飛びで取っていくニッチ分割型組み合わせの二通りを考える。
ここ数日このファイル上で思考を纏めているため、一日当りの分量が大変なことになっている…。
add TODO stack:
+ モデルの実装(6月中完了を目処に)
午前−午後と論文を読んだり研究に関して考えたりとか。午後増設用のメモリを買いにヨドバシに。512Mの最安が7000円強と高めなので別の店も見て回る。結局バルクで5000円弱のものが置いてあったソフトアイランドにて2枚購入。
夕刻より共用端末のリプレースとか色々。いろいろとトラブルはあったけどまあ使えるようになりました、と。それより計算機が一台死亡していて電源が入らない。一年間保証が12日までみたいなので早いとこ修理に出さないと。
明日6/10(火)14時から研究についての相談(第二回)とのアポイントを取る。
つまりそれまでに現時点での状況を文書化する必要がある、ということ。モデルについての基本的な構成は一段落しているので、下記の問題点についてもあらましを纏めて(下記のままだが)持っていって併せて相談してみることにしよう。
この前の研究計画に対して返答がくる。
メールによる研究の話し合いは意外に良いかもしれない。相手と向き合って議論できないというデメリットはあるものの、僕のように思考遅延がある人間には相手の意見を反芻できる点で文書でのやり取りは望ましい。少なくとも僕の性分には合っているような気がする。理想を言えばチャット(or筆談)の形が一番良いように思うが、相手との制約もあるし難しいか。
それはさておき、以下の問題点についてどう対応するかについて考えよとのお達し。
相談に言った結果と上記の考えを進捗状況としてまとめて提出する。週末、13(金)まで。
[ECOLOGICAL MODELLING 3rd ed. 適当にサマライズ]とstack listに書いてあるのだが一向に進まない。三中さんに倣って登攀記録でも付けようかしらん。
add TODO stack:
+ 進捗レポート提出(6/13まで)
午前中少しだけ論文を眺める。
先日借りた論文は半分くらいが当たり、というか結構重要な論文であるようだ、という程度に眺めただけ。後日きちんと読むことにする。
猪苗代湖を見に行く。ただし車。
仙台発→R4南下→福島→R115→猪苗代湖一周→R115→福島→R4北上→仙台。380Km/11h。
いろいろ降りたり歩き回ったりしてるので距離の割に時間がかかっている。猪苗代湖を一周するのに南西部が山道(一応舗装はされていた)で落石等もそのままになっていた。金属製の防護柵がぺしゃんこになっていたりと結構怖い。
帰路R115では道の駅つちゆ付近で霧が出て視界約50mくらいまで濃くなって大変だった。霧道は初めてなので良い体験ではあったができれは2度と御免だ。
本当は裏磐梯の湖が何個かある辺りも行きたかったのだが、有料道路の上に時間もなかったので断念。面白そうだったのだが恨むべくは金欠である。
そちらに行かなかったおかげか21時前に帰宅。
午前中仕事、午後私用、夕刻仕事。徹夜明けでこれなのでそのまま帰宅して寝ました。
午後起床。仕事のある日の出勤時間くらいに寝てるのでこうなる。そろそろ元の周期に戻さないと。夕刻輪読回。
昨日の続き。基本式ごとにちょっとした解説等を書き込み。意外と時間が掛かるなあ。今日はコレでおしまい。トレードオフなどについては論文を持ち帰る、けど、読まないような気もする。
午前中セミナー、午後勉強会。
よく考えたら6日(金)は旧生物学科あげてのソフトボールであった。僕は参加しないが何かと都合が悪いのではなかろうか。ということで自分内期限を9日(月)に延ばして、そのかわりに、すぐにでも作業に移れるようにアイデアを纏めておくことにしよう。
同じ研究室で個体の分散の進化についての研究をしている方に、移動能力と局所競争能力にトレードオフをかけたモデルを使っている論文はないだろうか尋ねたところ、すぐに4・5本纏めて持ってきてくれた。ありがたい。当たりがあると良いなあ。
個体ベースの方の基本式の構成を書き上げ。あらましを書いたところで力尽きる。Droopのモデルの基本理念を思い出すのに一番時間が掛かるというのがなんとも情けない。まだ一部アヤフヤなところが残っていそうなので一通り追いかけておくことにしないと。明日中には終わるだろうから、これが終わればトレードオフの方を詰める作業に掛かる、と。
実のところ実際の実験に使う個体群の形質セットの検討が最大の難関のような気がしてきた。どうしたものか。さしあたりAndersen(1997)の本と睨めっこかな。
教官殿と打ち合わせ。が、尋ねてみたところ打ち合わせの資料をまだ読んでいるとのことで、今日中の話にはなるまい(→なりませんでした)。次までに新しい資料を用意するくらいのペースで進めるのが良いペースだろう。
一日中論文を読む。途中雑用が入ったり仮眠をとったり。少しペースが落ちているが、週末に余暇は取ったので気力自体は落ちていない。少しペースをあげよう。
基本に立ち返る意味でGrover(2002), J.thor.Biol.(2002),214,599-618 を読んでいるのだが、こんなに面倒なモデルだったっけ? 整理能力が落ちているのだろうか。それとも基本ができていないからそう感じるのだろうか。後者だろう。
基本はMichaelis-MentenをLiebigで2資源に拡張したモデルのはず、という我ながら無茶苦茶乱暴な理解である。分かりやすく纏めておかないと必ずや破綻のときが訪れるだろうというわけで、現在纏める作業を行っているのだがやはり面倒だ。
Groverの一連の論文を読んで纏めたらトレードオフに関するまとめ。それが終われば植物プランクトンと動物プランクトンの形質セットについての検討。なのだがGroverの論文はどれもこれも長い、ひたすら長い。
モデルの詳細を詰めるのが自分の目標期限で今週中。…あと三日? こちらを急ぐべきだ。6日(金曜日)にアポイントを取りに行くことから逆算すると、明日中にGroverのモデルを整理、明後日にはトレードオフについてアイデアを出しておくというペースになる。好いペースだが纏めきれるかが問題だ。
土日は一日中部屋にこもって延々と本を読んでいた。寝るか読むかの生活はシヤワセの原点です、とかなんとか。晴走雨読を目指したいものだ。
明日の状況報告に向けて資料の作成、というより前回モデルの相談に持っていた分に追加と手直し。とはいえ、上半期の研究スケジュールプランまで含めるとかなりのボリュームではある。
生態モデリング学会のメーリングリストに参加申請。そういえば応用生態工学会の方のMLは寂れたのだろうか。自己紹介が数件とアナウンス以外に一向にメールが流れない。出だしから転けてしまった感がある…
トレードオフについて。
多くの動物と植物の群集で多種共存を説明するために、多パッチ多種系モデルにおいて、"a superior competitor has a lower colonization rate or a higher extinction rate"というtrade-offを仮定するのは一般的な方法らしい。( Kondh,M.(2000), Proc. R. Soc. Lond. B(2000) 268, pp269-271 )
トレードオフの掛け方についてはもう少し検討しないといけないが、文献についての目処は立ったので基本は[競争能力−死亡率]と[競争能力−移動率]で掛ける事になりそうな感じ。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}