80 論文+モデル:空間パターン成因モデルのまとめ
50 モデル:空間介在2種競争のCNRモデル改⇒(ASS or PoEからのアプローチ?)
40 論文原稿直し:CNRによる競争排除遅延モデル
30 モデル:侵入による多様性増加プロセス
-5 共食いモデル
昼過ぎ起き。
ちょい前に送られてきてたのをメンドくて放置してたんだが、いい加減締め切り間近なんでお仕事して再送。商業主義というのか周辺含めてプロっぽい、偏見か。夕刻まで。
ミジンコの空間的行動特性から藻類多様性を考える(仮題)、つづき。
文献を見つつ、モデルのデザイン修正。浅いところにユスリカ、…ねえ。まあ魚みたいのを仮定するよりは当初のデザインどおりstatic environmentではあるから、餌を追っかけるパターンとのコントラストにはなりそうな感じではあるけど。どーにも。
ちょっと手間が嵩むけど、いっそのことpredatorもカッチリとIBMでデザインしてしまおうか? いまのgrazerのモジュールを流用して、摂餌のところだけ改造すれば良いような気もする。
共食いモデル。
小人閑居したんで、とりあえず思いついたことを取りとめも整理も無く書き留めておく。
食物網(栄養段階)の進化・発達を考えると、系全体のエントロピー自体は大きくなっていく。しかし、その中のある循環をピックアップするとそのシステムの進化は収束系(のはず)である。なぜなら、転換効率の観点から考えると、たとえばFagan and Dennoがいうように、近接する栄養段階やintra-guildを摂餌するほうが獲得/努力の効率が高くなる。なので、その方が自身に有利のはずだからである。
しかし実際に「一般化された生態系」では、共食いやintra-guild predationに較べれば、grazer・omnivoreの方が普及しているように見える。だとすると、栄養/エネルギー転換効率的観点から見た場合に、omnivoreで進化が止まる理由は何だろうか。grazerを栄養段階の下部に留めておくconstraintは何だろうか。皆が皆、草食わなくなったら誰も生産者からのエネルギーの運び手がいなくなるから、という説明は成立できない。草を食うからこそ有利に事情が必要となるはずだ。
この説明として、ひとつには、餌と自身の体構成が完全にバランスマッチしてしまうとエネルギー(炭素)に制約が掛かってしまうというのが挙げられる。肉(N・P)だけでなくご飯(C)も食べた方が日常生活でのmaterial lossが小さく効率が良い、という理屈だ。
この説明を当面受け入れるとして、栄養段階の進化において、random feederがfeeding selectivityの突然変異によって、自身のenergy/materal efficiencyに近いものへの選択性を上げていくというプロセスを考える。このプロセスによって、生態系内にはpredatorにとどまらずintra-guild predatorまで進化するものもいれば、omnivoreが最適であるとするものもいるし、やはり草しか食わないものがいる。grazerがgrazerのままであるというのは、草食いからの進化には消化系の改善が必要になるというコストによって、もしかすると説明できるかもしれない。では、まだ収束していないというアイデアはとりあえず措くとして、predatorとomnivore両者の進化分岐条件は何だろうか?
とりあえず、case by caseというのは禁止で。
自宅作業。
ここしばらく取り留めも節操もなく文献を読み漁ってたんで、いい加減整理しないと何に関しての論文なのかわからなくなりそう。というわけで、とりあえずジャンルと優先順に整理、結構時間がかかる。
午後過ぎから私用で外出。翌明け方まで。
早朝起き。
本を読んでから研究室。
『予言の守護者 』(デヴィッド・エディングス)、読了。
ベルガリアード物語と題した長編ファンタジーの一巻、復刻版。読み始めた当初は、ありきたりな探索物だなと感じつつページを進めていたのだが、一巻を最後まで読み終えて脱帽。これは凄い。
素直な読み方をすれば単に『指輪物語』の焼き直しに思える。王道ゆえの面白さはあっても、多少なりともファンタジー作品に触れたことがあれば斬新さを感じることは多分無いだろう。しかし違う、この作品の本質は多分違う。ありきたりとか言う生半可なものではない。これは「教科書」だ。そういう視点で読むとこの本がどれだけ凄いか判る。背景設定から登場人物、細かいイベントにいたるまでの全てが定型フォーマット・テンプレートであり、どれひとつとしてその枠から絶対に逸脱しない。でも面白い。
嫌味や褒め殺しのつもりで書いているわけではない。会話による緩衝材はあるものの、ここまで一切逸脱しないままきっちりと詰め込まれているのは、膨大なデータベースと緻密な計算なしでは絶対にありえない。普通の作家ならキャラクターやストーリをいじりたくなる(と、思う)が、崩さないからこそ完成された作品を作り上げている。オリジナリティなど必要ない、テンプレートの組み合わせだけで一級のエンターテイメントは成立する、という著者の信念が垣間見えるように思える。この作品をフィルターとして通すことで、他の作品を読むときに、そのオリジナリティやアイデアの独自性だけが浮かび上がることになるだろう。
と、これだけ書いておいて、2巻以降がただの焼き増し作品になってたらどうしよう。僕の穿ち過ぎな読み方ではないと思うんだけど。
朝、研究室。
昨日仕掛けて帰った計算結果の確認。ばっちし予測通りですな、よしよし。正午前、私用で町へ外出。暑い。
夕刻、用事のついでにトガシ君のところに先日の結末を聞く。とりあえず別のアプローチからデータ解析を試みるとのことで、晴れて御役御免。
論文原稿修正つづき。
引用文献の書き漏らしと情報ミスが無いかのチェックと整理。アタマを使わない作業ではあるが案外シンドイ。
侵入モデルつづき。
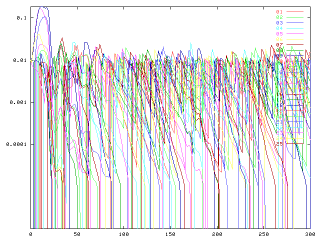
ロジックがモデルの上で曲がりなりにも成立するのは確認できた、と。これでひとまずは、絶滅実験だろうが侵入実験だろうがCNRを考慮したほうが藻類多様性が大きくなる、というのを自分の中で疑わないですむ。多種系ならばという限定付きで。
問題は幾つかあって、結局のところ、残る種数は移入と絶滅のバランスに依存してるので、移入が多すぎれば問答無用で飽和するし少なすぎれば一種だけになる点。さらにテストのときのように移入量が多いときには、マスが同調しながら多種⇒少種の時間振動が起きるという別のロジックが成立する点。後者はちょっと厄介だけど、同様のロジックは誰でも簡単に思いつく筈。おそらくは"paradox of enrichment"に関する周辺論文で議論があるんじゃなかろうか? 探すとしよう。
あとは条件範囲とレプリケートをとるために裏で計算させつつ、とりあえずこのモデルに関してはカットオフ。
catastrophic shiftモデル案のつづき。
現時点で一番モチベーションが高くて一番まとめたいアイデアにもかかわらずまとまらない。空転してるだけで、何が面白くてどういうストーリーにできそうなのかが漠然としすぎなんだな。
朝起き。午後過ぎ、研究室。
朝から用事であちこちへ。陽気の所為で厚手のブルゾンでは汗ばむほど。暑さが辛くならない快適な時分の内に遠出などしたいところではある、が。
夕刻、カワタ先生よりMLが停まってないかとの問い合わせ。どうやら勘違いだったらしいのだが、肝の冷えるようなことをサラリといわれると心臓に悪い…。
カワタ先生と幾つか論文修正原稿について遣り取り。判らぬと削除されたのを書き直して復活させた箇所についてやはり判らぬという指摘。条件についての説明するのに留めて長々と解釈せんでよいということになる。納得してしまった時点で僕の敗け。指摘された箇所について説明した後で自分で原稿を見返したら、無駄が多い割に散漫で、説明した意図のとおりには書けていない。
もう一点の指摘についての議論の決着は先送り。
侵入モデルつづき。
昨日のうちに仕掛けておいた結果が出ているので眺めてみる。CNRなしの条件下では、条件中でもっとも有利なやつが一旦居着いてしまうと、侵入しても競争に負けて絶滅してしまう。よって、時間的には強い奴がでかいマスを占めてて、小さいアバンダンスでの種の入れ替わりがチョコチョコとおきている。ここまではロジック通りで気分が良いくらい予測のまんまだったのだが。
対して、リサイクルによる栄養塩量・比の変動が作られる条件下では、確かに侵入も容易になっている。ここまでは持論でのロジック通り、Andersen(1997)のような排除則の強化には必ずしも繋がらない、ここまではいい。だが、すべての条件で捕食−被食系の大きな振動が起きている。排除モデルの方では振動が見られなかったような低い栄養塩供給量の条件下でも起きている。結果として、侵入しやすいが皆で増えてまとめて排除されるという状況。
何が原因なのかをあれこれ考えていたら暫くして理由に気がついた。移入によって系全体の被食者バイオマスが潜在的には線形増加できるのが原因。このこと自体は定期的な移入を前提としているため回避できない。
テストで仕掛けた条件では、流入してくる藻類のバイオマス(かける頻度)が多すぎて、供給栄養量にかかわらず被食者の炭素量バイオマス(に伴う含有N・P)が一気に富栄養化しているわけだ。食われるCが増えれば排出されるN・Pも絶対量で多くなるので、リサイクルとして反映される条件下では上述の結果になった、と。
CNRの無い条件下で、同量の流入があっても大きな振動が起きないのも、傾向としては排除モデルのときにかなり富栄養にしたときとよく似ている。
この程度のことはあらかじめ予測できた筈なのにウッカリしていた。やはり適当な数値で計算させるとこういう弊害が付きまとう。というわけで入ってくるバイオマスを目いっぱい下げてやり直し。予測としては、侵入と絶滅でバランスがとれて振動平衡みたいなのが出てくるか、もしくは、Andersen(1997)のいうとおりにN/P比の極端化が進んでやはり一種になるか。僕としては後者だとオモシロくない。
ところで、無機塩類の負荷量の増加でなく、生物量そのものの負荷量(?)の増大による富栄養化というのは、実際に野外でありえるんだろうか。そもそもが亜閉鎖系を想定した研究では想定外という気もするが、セストンの流入量に関するモデルは結構あるわけで十分にありそうにも思える。赤潮の流入による観察系の群集動態のコラプション…とか? 与太話だな。
ごく簡単に探してみたかぎりでは、Abrams and Roth(1994), Evol.Ecol.(8),2: 150-171.でimmigrationによってcarrying capacityが大きくなるといってるのがあるが、ダイレクトな議論をしている論文は無いようだ。
どうでも良いということでも無いんだろうが、定性的な議論としては無機塩の負荷による富栄養化と変わらない(内的増加が増えるか外的に増えるかの違いに過ぎない)からだろうか。
昨日のY字管、丁度用事があって置いてある部屋に行ったときに手にとって見たら全く似てなかった…。うーん、勘違い。
早朝起き。朝、研究室。
しばらく本を読んでから研究室。作業続き。午後バイト。
侵入モデルつづき。
もそもそと手を動かす。何も居ないとこに逐次撒く形にしていたら、最初に藻類が増えるまでミジンコを支えられないということに思い至る。というわけで、初期種数を何種類か用意してランダムな組み合わせからスタート。
で、夕刻過ぎにとりあえず完成。うはは、やり過ぎた。無茶な条件設定をして、毎日のように藻類がどっかんどっかん流入してくるの図。移入速度>>(越えられない壁)>>絶滅速度、という感じ。種数が飽和してるがな。
…さて。実際に現実的に「ありそう」なパラメータを探すとして、野外ではどの程度の流入出フラックスがあるものだろうか? うまいこと適当な文献が見つかると嬉しいのだが。似たよなことをやっているHuisman et al.(2001)では、"… once every 50 days a new species was added with a low abundance of 0.1."としか書かれてなくて、実に適当に入れてみましたという感じになっている(まあ、この論文では、パラメータの設定全般にわたって、定性的なものが見たいんで野外での現実性とかはとりあえず棚上げしとく、みたいな言い訳が書いてあるんだが…)。
ん、Huisman et al.(2001)の真似をしないで、移入「率」として一日あたりの確率判定をさせたほうがよいのか? 本質的な所では無いんだけど、完全なパルスで入ってくるよりはそれっぽいのかもしれない。あとで試してみるか。
とりあえず適当な数値で侵入モデルの計算を流しつつ文献探しなど。夜、撤収。
朝起き、自宅作業。
先日再構成した自宅のマシンが予告なしでフリーズする現象が多発する。どうやら「マルチメディア」カテゴリのデバイス同士が衝突している感じなのだが…。とりあえず当分使わないであろうデバイスを物理的にはずして再インストール。W2Kの大容量HDDの壁に対しても、今回は前回作った大容量対応用の.regファイルが残っているんでさっさと処置。念のために中身をメモとして残しておく。136G以上の大容量ディスクをW2Kが認識できないときには、下記のの4行をコピーして.regファイルにしてからレジストリを更新する、と。もちろんregeditで書き換えても同じだけど。
Windows Registry Editor Version 5.00
[HKEY_LOCAL_MACHINE\SYSTEM\CurrentControlSet\Services\atapi\Parameters]
"EnableBigLba"=dword:00000001
必要なデータやアプリケーションなんかはその殆どが別ドライブからロードする構成にしてあるんで、こういうときのリカバリはあっというまに終わる。…と、言いつつ朝起きてから始めて昼過ぎ。
午後過ぎ、研究室。
これまでの経験上、こんなのやりますーというのだけ持って行っても「やれば」or「何がおもろいワケ?」の二択になるだけで実りのある話にはならん(かった)と思い返して、モデル書きのつづき。実際のところ、仮に僕に同じ相談されても、碌に纏められてもいないモデルのアイデアだけ持って来て、良いアドバイスというのを求められても困るだろう。面白そうなアイデアとか当人が乗り気ならやってみればというだろうし、そうでなければドコが面白いのかと聞き返す。野外やラボの実験計画だとコストもあるし、まさかこう適当には行かんだろうけども。
侵入モデルつづき。
ひとまず結果を出して見通し込みで、ここまでやりましたーという話のほうが進めやすかろう、という訳で、実装できる程度にはアイデアが纏まっている侵入モデルからさっさとコーディング。ASSモデルのほうもなるべく早めに纏めて一緒に持って行きたい所だけども。
侵入モデルのほうは修正した原稿の排除遅延モデルと対になっているというか、切り口を変えているだけであるけど、僕としては押さえておきたい話。侵入のパターンはHuissman et al.(2001), Ecol.letters(4),5:408-411.から拝借。
なんとか今日明日くらいには終わりそうな目処が立つ。…バグさえ来なければ、…れば。どっちかというとレプリケート数をいーぱい採る必要があるし、計算機がガラ空きな今のうちに、まずこれを片付けるのが正解だろう。
ま、モデルの話は兎も角、総体的なプランについてはどのみちキチンと話をせにゃならんのだけども。
catastrophic shiftモデル案のつづき。
Scheffer本みながら、なにか繋がりそうで繋がらないのがもどかしい。もやもやと漠然としたアイデアイメージからどうにも固まらない。紙に落書きの絵なぞ書いている。
大阪のときに雑談した内容で繋がりそうなものを、今頃になって思い出したんでメモしとく。ドラスティックなシフトの例。アイデアとしては、2-2-2なモデルになるのかしらん。拮抗的な関係にあった消費者のうちの片方が絶滅だか激減したせいで、珪藻がいなくなって藍藻が大繁殖した(逆かも)。どこの話で誰から聞いたんだっけ? これだけ聞くとgrazing selectivityの違いでオシマイ、という感じだが。塩害か何かによる環境変化がトリガーだった気がするんだけど、あー、きちんとメモしておくんだった。
ミジンコの空間的行動特性から藻類多様性を考える(仮題)。
相変わらずセンスというかヒネリがないと言うか。行動特性⇒空間パターン、と、空間構造の多様性への効果、が二本柱。といいつつpredator avoidanceとfood aggregation(あとランダム)しか考えてないけど、ざっと論文漁りをしてみた感じでもどっちかの話であることが殆どなんで、ま、とりあえず良しとしておこう。
話の組み立てのための論文探し。空間構造と競争と多様性の話はAmarasekare(2003),Ecol,Letters(6):1109-1122.を読み返し中。適宜ここから孫引きしていけばよいか。
行動特性⇒空間パターンのほうは、大阪ではFolt and Burns(1999)のTrEEのREVIEWを引いたけど、パターニングとスケールの関係性を何度も強調しているので、これをこのままスケール的な話をとりあえず棚上げしているモデルに引くのは躊躇われる(議論の方では避けられないだろーけども)。
で、行動特性と空間パターン(verticalでないやつ)を他に探す。horizontalな話だと、みんな「けみかるきゅー」が好きすきーという印象。他に無いんかいと云うくらいに。新しメなところで、Roozen and Lurling(2001), J. Plankton.Res(23),8:797-808.とVan de Meutter et al.(2005), Oecologia (142): 489-499.の二つを眺めてみる。前者はbehavial responceの話で直接空間パターンがどうこうと言ってる中身ではないけども、まあまあ良く纏まってるんで取り敢えず。本題と関係ないけどRoozen and Lurlingにはどこかで見たようなY字管が出てきて、これ似たようなのがカワタ研にも何故だかあるんだよね。文献を追いかけていくとVan Donkと仕事をしてる人たちらしい。
そういえばchemical cueといえば、反応拡散モデルを拡張した、ガチンコな行動応答の空間計算シミュレーションモデルを作ってた文献を見たような気がするな。そこまで細かいところまで突っ込むつもりは無いけど、何だったかな。このテのは多分Ecol.Model.かJ.Theol.Biol.だと思うけど。
horizontal pattern formationな話も探してみると上記の以外にも出てくる。Davies(1985), HYDROBIOLOGIA (120),2:103-105.とか。しかもDiurnal migration? ちょっと古いけど図書館にあったはず、複写に行こう。
あちこちから悲鳴のメールがいっぱい来ている。土日どころか昼夜もないお仕事の方々、お疲れ様です…。
朝起き、自宅作業。
無印のときとは放送局(MBS⇒TBC)が変わったけど2週遅れで仙台でもジェネシス開始。一話目から燃える。日曜日の朝はさらに前倒しになってしまった。
Sterner and Elserの化学量本のCNRのトコとか他に持ち帰った本の消費者関係をぱらぱらと流し読み。consumerによるカスケード効果を考慮した藍藻と緑藻のcatastrophic regimeな競争モデルがあったと思ったんだけど見つからず、SchefferのASS本ではなかったっけ。その後の論文だったかな。
夕刻、金曜日の統計の話に関してマツシマさんからメールをいただく。いくつかアドバイスをもらったんで早速試してみるとしよう。意図的に避けてきたRにも結局触る羽目になりそうな感じ。
ちょっと遅めに起きてから山開きした蔵王に行って午後過ぎに研究室、…の予定だったのだが。
龍泉洞。
友人数名に連れられて岩手に。東北道(仙台ー盛岡)〜R455(盛岡ー岩泉)。盛岡で昼食後、午後過ぎに龍泉洞着。鍾乳洞内の地底湖。土壌濾過水が集まっているために透明度が非常に高く、(たぶんセッキ)深度40m強という話だった。
ついでにリアス式海岸を見ようという話になって、帰路、R455(岩泉)〜R45(宮古ー釜石)〜R263(遠野ー北上)〜東北自動車道(北上江釣子-仙台宮城)。総距離700Km、日付が変わる頃自宅着。
早朝起き。
午前中お仕事。午後、研究室。
モデルについてあれこれ。…脇道に逸れて共食いモデルに関してあれこれ思索。material-materialベースでのefficiencyをあれこれ検討してみるけど、面白い案が浮かばず。生食循環に組み込んでどう動くか、…イマイチ。
午後過ぎ、輪読会(占)。今の本が終わった後は教科書でなくてゼミ形式の論文紹介になるとか。ま、それはそれで。
トガシ君データ解析続き。これも思った以上に長引いているなあ。彼の顔を見るまで毎回忘れているのが原因、という気もするけども。というわけで輪読会の後でとりあえず(色々な問題点をはらみつつ)Iδを出すだけ出してみる。すべてのデータで集中分布であるという結果に。最初は計算結果が違うのかと思って、ランダマイズデータや一様分布によるデータで検算してみたのだがそれっぽいF値になる。
自前の理解が間違っているかもしれないので、解析が正しいのかどこかに問題があるのか判断できず、とりあえず保留。が、検定時に(とりあえずで)仮定した前提に問題があるような気がする。
夕刻、昨日送られてきたカニをマサヤ君が食べているのを眺めていたら、「良質のタンパクですよ」なる台詞。最適餌質摂餌なキャラクターという認識なのか。昨日のうちに生きているのを散々弄繰り回して造形を楽しんで満足していたので、「斯様にhandling timeの大きなものは選択しない。」と答えたらわざわざ身だけ寄越してくれた。旨かった。
夜、頼まれて管理だけしている某MLのadminあて(システムに関する要望と苦情の受付アドレス、登録等のコマンドの受付とは別)に大量の件名・本文なしのメールが殺到する。何事かと思っていたのだが、事の真相はMLユーザの母集団でガイダンス中にMLのadmin宛に何も書かないメールを送信すると登録されるという説明があったらしい。
そういえば、確かにずいぶん前にadmin宛に間違えて送信してきた人がいて、間違えていましたが今回は登録しましたという旨の返信をした記憶がある。それが間違って学習された模様。
流石にいちいち手作業で対処するのは冗談ではないので、説明しなおしてもらうように案内を送る(システムから送られてくるレポートを読むに解決されたらしい)。教訓、情けは人のためにはなるかもしれないが、自分のためにならない(ことがある)。妙な仏心を起こさないで機械的に処理してしまえば自分の手間は増えないかもしれない…。
早朝起き。
『人間の本性を考える 心は「空白の石版」か (上)』(スティーブン・ピンカー)、ようやく読了。中巻を注文。いくつか思うところもあるが最後まで読んでから。下巻まで読み終えられたら感想など。
自宅作業、侵入モデルのほうの続きを少し。昼、散髪のち、研究室。
午後過ぎ、研究室。
論文修正原稿、最後の部分にいて少し話す。ついでに、今年度のプランについての話し合いを週明けにアポイント。
昨日のディスカッションの最後の部分は、話をして程度問題に過ぎないという結論に落ち着いたので削除、リファレンスの体裁については放置。メールでカワタ先生に提出。
そろそろまた後手後手のパターンになりつつある。このまま行くと例年通り自滅パターンなので、生態学会のポスターに使ったモデルの整理しなおし。成因としてしょーもないものを全部落として、top-down、bottom-up、ランダムの3つに切り分け。top-downについての仮定というか想定がどうにもアヤシイのをどうするかが当面の解決課題。どちらにしてもこの部分はモデルを作り直す必要がある。今回の目標は日本語の下書きを書かないのと時間を掛けないこと。
うーん、新しいモデルのほうのアイデアが整理されないまま後回しになっていく…。
朝、研究室。
午前中セミナー(カ)。前半担当は福原君で卒論研究の紹介。アリマキに随伴するアリが宿主植物の成長やアリマキ捕食者に対してどのような影響があるかを調べた内容。農学部での研究らしく最後に応用への方向性についての議論をしていたが、バンカープランツという概念は知らなかったので勉強になった。簡単な理解ではapparent competitionを応用することで作物への害虫を抑制しようというものらしい。調べてみると既に商売としても成立しているようだ。
後半担当は吉野君で論文紹介。Kingston and Rossiter (2004), "Harmonic-hopping in Wallacea's bats", Nature(429):pp654-657.の紹介でコウモリの声の周波数の違いによって繁殖隔離⇒種分化のメカニズムを示した内容。早口気味だったけどよく整理されていて面白かった。周波数の違いが生じる機械的な要因について質問したが不明とのことだった、ラザロさんが後から教えてくれたところによると、コウモリは聞こえ方の違いに合わせて(自分が聞こえるような)周波数を発声するらしく、聴こえ方が聴覚受容体のニューロンの数で決まり、その数が遺伝子によって調節されている、というメカニズムらしい。
午後、修理から戻ってきた計算機を一台潰して、山口さんに計算機のセットアップ手順を覚えてもらう。津田君が熱心に教えてくれたので、僕はいくつかヘルプを出した以外には特にやることなし(実に素晴らしい!)。結局3回くらい入れ直しやらなにやらやっていたらしく夜遅くまでやっていた。午後過ぎ、私用で外出。
論文直し、ひとまず完了。結局イントロダクションのパラグラフを丸ごと書き換えたりとか、大幅改修。ディスカッションの一番最後は、少しカワタ先生と話をすることにする。
午前中、研究室。
論文直し続き、インターバル込みで夜過ぎまで。
議論部(再)。先日、カワタ先生に『ホントに微小時間で空間的なリサイクルの差異が効いてないというのが確かめられていない』という指摘をされてその場で反論できなかったのだが、よくよく考えたらそのためにリサイクル直後の環境の栄養塩状態を1ステップのオーダーでモニタしたんだった。その旨書き足し。
もう一点の初期値依存性についての反論、というか言い訳は、書いてはみたけどかなり苦しいような。追加というか補足的には、いま書いている(途中の)逐次侵入が起こり続けるケースを想定したモデルで確かめるという形になるとは思うのだけど、…同時競争開始のみで説得するような良いアプローチが思い浮かばない。この点はもう少し詰めて考える必要がある。
議論部分の総括に関する修正案に対して、重要でないと削除されている項目に関して反論。その他の箇所は考えが纏まらないのでちょっと放置。
イントロダクション直し。これまた真っ赤の上、一部に構成再考の指示。半分まで進捗。が、こちらも一番の壁は持ち越し。
午後過ぎバイト。
天文学か何かを受講している学生さんから課題に関する相談。半年振りくらいのまともな…と思ったのだが。課題内容は黒体輻射に関する作図と数値計算(だと思われる内容)。もしかすると物理か何かでまったく別物かもしれない。
問題は、学生さんがプログラムはおろか作図その他の操作から全く分からない、のみならず、課題に関しても数値計算の手法の指示があるだけでその手法がどのようなものかについては全く説明がないという点。
聞いてみると受講者の大方は数値計算はまるきり分からないはずとのこと。その学生さんたち(二人)が特別アレゲなのかとも思ったのだが、黒体輻射の理念とかプランクの式についてはきわめて明快に説明できるようだし。仕方がないので一通り簡単にレクチャしたのだが、いったいどういう授業内容なんだろう。
で。研究室に戻ってその場にいた数人にその話をする。簡単のために「われわれで例えるとLotka-Volterraモデルの個体群動態を数値計算して結果を作図して提出しなさい、というようなものかな」と例えたところ、「ろろら・ほにゃらら?、とやらは種名かなにかでしょうか?」なる返答が返ってきて吃驚した(僕も似たようなことはよくやるけど…)。
その場でLVモデルとはなんぞやというレクチャになる。それで良いのか生態システム専攻のM2とM1とB4(進学希望)の各一名。M2氏は「何かそんなのがあった」と思い出したようだったけども。生理方面だけで受験できる生命科学研究科の院試の弊害が此処に。僕も相当に不勉強なんであんまし強くはいえないけども、LVモデルくらいは学部の授業で勉強した記憶があるような。
とここまで書いて、普段僕が発する各方面での頓珍漢な内容の放言は、おそらく周りには斯様に聞こえたり、そんなことも知らぬのかと吃驚されたりしているのだろうなあと思いかえす。これまで僕が系統解析とか統計とか安定同位対比とか、その他諸々さまざまな事柄に関して無知蒙昧をさらけ出してきたのとまったく何も変わらないということだ。
まあ、ちょうど輪読会で『An Illustrated Guide to Theoretical Ecology』(Ted J. Case)をやることだし、そこらの知識は十分に補充されるであろうて。
午前中、自宅作業。
昨日からのリカバリの続き。アプリケーションの設定などについて、自宅でも使っているものを抽出したりアーカイブして転送したり。
午後、研究室。
とりあえず、作業環境はほぼ回復。が、来年以降の研究計画のために集めていた文献がトンで、これをまた集めなおすのが億劫だ。
午後過ぎ、セミナー(マ)。
前半担当は今回もトップバッターということでウラベさん。今回は学生に求めることについての話はごくごく軽めに流して、大半は研究紹介の話。デトリタス食と摂餌選択性を考慮した植食者の共存モデルに作りましたという話。かなり前にデトリタスに着目すると競争がどうなるかという議論をしたことがあったけど、多分当時すでにこのモデルが念頭にあったのかな。
LVモデルをシンプルに拡張した、判り易くかつ言えることの沢山ある良いモデル。現実に観察されるパターンとの対応についての質問に対しては「パラメータ次第」という答えが返ってきた。まあ予想通りの返答だが、ある観測事実を説明するモデルというよりは仮説を説明するタイプのモデルなのでこれは仕方がない。「海のCopepodのペレットがコーティングされるのは、もしかすると競争的な意味があるのではないか」という議論は面白かった。現実にみられる「不思議なこと」に対してアイデアを提示できるのはとてもよい。
要旨には、時間があれば山の話もしますとあったけど、前半のイントロダクションをかなり丁寧にしたため時間オーバーでその話には触れず。珍しくカワタ先生が居たのは、もしかするとホントはこっちの話を聴きに来てたのかも。
後半担当はマキノさんで「底プロ」進捗報告。あちこちからコペをとってきて遺伝子を採って比較したところまで。チバさんが指摘していたところによると種内の変異が驚異的に大きいらしいのだが、そちらの方面に親和性がないのもあって、そういうものかというくらいの感じしか受けなかった。結果についての仮説考察なども提示してくれれば絡みようもあったのだけど。
先日の話、Langtonでなくて、マトゥラーナとヴァレラの『オートポイエーシス―生命システムとはなにか』だった。確か人工生命やりたいなーと思ってたころに買った本で、かなり徹底した還元論というか機械論者による著書。生気論で言うところの生命の合目的性についてもあれこれと述べてた、と思うんだけど、えーと。…もう一度読み返すことにする。
でまあ、定義と認知の問題というのはあながち外してないとは思うけども、それを言ったら議論にならんので、この辺の(疎覚えな)話を引き合いに出しつつ、こういう見方の人が居て別の見方の人も居てという各論紹介的に軽く話す。
夕刻、フクハラ君のセミナー要旨を眺めていて、餌転換効率だのギルド内捕食だのというとてもとても気になるキーワードを見かけて、(たぶん発表と関係ないだろうけど)質問。
餌転換効率の方は、どの餌で育ちがいいか成長率の違いを見ているだけという話だったんで、やや期待はずれ。テントウムシのギルド内捕食の話の方は、餌(アブラムシ)が短期間に沢山ある状況(草本)だとそれを専食する種がいるのに対して、木本ではアリマキを食べる他の競争者が沢山いて、その競争に勝つために競争能力を高めるついでに競争相手を食べることで利益を上げる、という話だそうな。
いまのとこ共食いモデルに何かしら反映できるアイデアは直ぐに思いつかないけど、頭の隅のほうに覚えておこう。
夜、論文直し続き(再)。土曜作業分くらいは挽回。二度目の分だけ質が上がっている、と思うことにしよう。
データの自動バックアップ。
ffftpを使って更新分だけ手作業でバックアップしていたが、手作業だと更新頻度が落ちてよろしくないという学習をした。というわけで、フリーで更新差分をインデックス化してくれてスケジュール機能を持つものを探してみると、意外に少ない。それでもBunBackupというソフトが見つかった。
物理的に壊れるのが問題なので論理パーティションでは駄目。自宅のPCのように複数のHDDを積んでいれば別のHDDに逃がせばよいのだがノートだとそうもいかない。というわけで、smbで共有したネットワーク越しのマシンにドライブレターを振って、とりあえず3時間おきくらいで自動的にバックアップ。お、結構使いやすい。
バッドラックな日というのがあるようで、布団を干せば雨が降り、原稿を書けばHDDがクラッシュし、HDDを買いに出かければ背筋を痛め、と、終日何かやるたびにトラブルが付いて回る一日だった。挙句、自宅に帰る前に買った弁当をその場でひっくり返すという落ちまでついた。逐一さえないコント調なのがいっそう物悲しい。
午前中、研究室。
午後過ぎまで原稿直し。昨日土曜日からの続きで議論部に目処をつけ、イントロダクションに着手、の途中でフリーズし、HDDから異音。またか。またなのか。一年も持たずにHDDがクラッシュ(物理的に)。で、土日分の原稿がロールバック。とはいえ、そのままでは支障が出るので、データのサルベージはすっぱりと諦めて、さっさとHDDを買ってきて環境の再構築。夜まで掛かる。予定外の出費、7500円。
今回の教訓。一日おきといわず、半日くらいおきに『自動で』別のマシンにデータを逃がす手段を模索すべきである。もうひとつは、USBフラッシュメモリを挿したままリカバリをしようとするとそちらに再インストールをはじめようとして失敗する。同じ容量のディスクを買ってきたのに容量が足りないといわれたのだが、最初は気が付かなかった。
なにより、ディスカッションの直しは結構きつかったのに、もう一回やりなおしというのが痛い。いやはや。
午前中、おしごと。午後、研究室
論文直しつづき。修正案を受け入れたりそれに沿って書き換えたり反論を書いたり。一番厄介なのが、各要因についての説明をどうやって説得するかがかなりの難問。カワタ先生からもそこをどうするかがキモだというコメントを戴く。
夕刻、本日は川内にて河田研と占部研合同のお花見。論文議論部なおしで煮詰まって途中で息抜きがてら合流、かなり遅刻。
到着してすぐにウラベさんから、コンピュータウイルスは生命といえるのか、なる問いかけ。一体なんでまたそんな話を?と訊くと、授業後に学生さんからの質問だったらしい。生命をどう定義するかによりけりでしょう、と言ってお茶を濁そうとしたのだけど流石にそんなにアマい相手な筈も無く、あれこれと議論。なんだかそのテの本を自分で持っていたような気がするんだが、何だったかな。GC LangtonのA-Life本に書いてあったかもと言うような話をして、気になったので研究室に戻ってきてから調べてみたけど、そういう話は載ってなく。人工生命か複雑系がらみの本で、その手の議論を扱った本があったんだが手元に無い。
一時間程でその場を辞して研究室に戻る。論文直しつづき。今週中(日曜日込み)で、何とか形にしたいところ。
午前中、研究室。
自宅でちょっと本を読んでから。あまりに好天快晴なので、やや遠回りをしてぐるっと廻ってから研究室へ。
午前中、昨日の課題となった箇所について、幾つか本を積んで勉強しなおし。昨日の不明な点に関して周辺も併せてほぼ理解した。とはいえ、さてこれを応用してデータの解析案を考えよ、というのにはまだまだ習熟度が届かない。Iδ駄目かも。マツシマさんに議論を吹っかけて無理っぽいのうというような流れになりつつ、あれこれ悩む。カワタ先生に質問してみたら、やはりIδはたぶん駄目でANOVA使ってみたらというアドバイス。ますます迷走中。
正午前から論文直し、夜過ぎまで。単順な英作文の修正に留まらないので悩ましい。文献を読んで考え直したり、文章を書き直したりして微速前進。
途中で気分転換のつもりでヤゴ空間分布解析ネタを考えつつ、息抜きになってないというか、むしろ深みに嵌りつつある…。
議論部、英作文以外も大幅に修正されていてなかなか進まず。四苦八苦しているうちにイントロダクションも帰って来て、構成を見直したほうが良いかも、とある。日本語版を書いてたときに散々敲かれて軟着陸した筈だったのに。…のに。議論部の修正と加筆を7割がた終えた所で、此処から先にカワタ先生による膨大な加筆案が待っているが、力尽きて撤収。
『An Illustrated Guide to Theoretical Ecology』(Ted J. Case)注文してしまった。前々から欲しかったのだけど、数年前から何回か候補に上がっていた本で、今年度の研究室輪読会の読本とのことでよい機会なので購入。うむ、年度始めの月から本代がエライことに…。
今年から博士課程については参加は任意という方針になったこともあって、パスかなーと思っていたのだけど、この本なら参加したい。しかし、どういう経緯でこれに決まったんだろう。と思って訊いたら、前の本は皆が猛反対したじゃないかとの由。そーだった。
正午前、研究室。
朝、冨樫君からデータが来たんでちゃちゃっとモデルに載せて解析。まあ予想通りにはなった、のだけど…。
午後過ぎ、計算機部屋のレイアウト変更。
棚を一つ増やして計算機の配置にゆとりを持たせる。この件に関してはツダ君が頑張ってくれて、僕はせいぜい注文をつける程度。もう計算機に関しては大体任せてしまって問題ないだろう。サーバのほうの引継ぎについてもぼちぼちと。新B4の山口さん(彼女は僕などよりも遥かに物の飲み込みが早く、あっという間にプログラミングは覚えてしまった)も割と積極的に覚えようという意欲を見せているので、二人して頑張ってもらうとしよう。
で、そのために作業内容全般に関してドキュメンテーションせよと津田君から言われてるわけなのだが、正直なところ、僕がこの研究室内で管理してるのは内外向けのMLサーバと研究室の計算機チーム用の統括サーバだけど、手作業をしない為の努力をたくさんしたおかげで作業の大半が自動化されているので、実はルーチンワークとしてやることは殆どない。書き起こすのがメドイというのもある、けども。
統括サーバはハードウェア構成等がやや特殊うえ、gateway他、いろいろなサービスをまとめて担っていて、…落ちたらどうしよう。大体、デバイスドライバをコードの一部を書き換えたりカーネルモジュールのレイアウトを弄ったり(修士に上がった頃はそんなことばかりやっていた)と、いろいろ遊んだ挙句動いている危ういマシンなので、もう一回同じ物を作れといわれても正直僕の手に余る。
MLサーバについては既に物理的に半二重化が図られているので、HDDがクラッシュした場合に若干ロールバックが起きる以上の障害はない(はず)。こっちはオチると学内のかなり重要なMLだけでなく、いつのまにか管理していた学外向けのMLとかも停まるんで、割と真面目にメンテとか障害対策がされている。こっちは30分もあれば別のマシンに転生できる。
上記の統括サーバにせよMLサーバにせよ、元々は自作マシンを改造/増築したところに色んなサービスを試験的に載っけていった物が、いつのまにか正規運用されているという現状がある。趣味のテスト機がそのまま転用されるというのも不思議な感じだが、当研究科は特にボランタリーな側面が色濃くて、生物系の情報インフラ自体もタジマさんとハシカワさんがいないと殆ど動かないといっても差し支えない運用体制になっている。ハシカワさんに至ってはタジマさんの後輩である以上の理由すらなくて、ですね。
まあ工学部とかの話を聞いても、大学のインフラ事情(のすくなくとも一端は)こんなものらしいのだが、情報インフラのホメオスタシスというのは、実のところ想像以上に綱渡り、という側面はあったりする。とゆか、自分が知ってる所は大半が個人で支えている。そういう意味ではカワタ研のように小規模でも後継者がきちんと育ちつつあるというのは恵まれてる、というのをオチにして、ドキュメンテーションは一部無理(自分でも動いてる理由がわからない)、…と云ったら怒られるだろうなあ。
午後過ぎ、カワタ先生より議論部が返ってくる。相変わらず真っ赤だが、今回は反論とか言い訳をさせてもらえる部分も結構ある、ので、前回のように英作文だけをひたすら直されるよりは凹まない。
夜、ウラベさんのところに。
冨樫くんからの下請けのヤゴ池分布ランダム性検定。出来たのはいいけど、どうにも違和感というか解析方法に釈然としないので、カワタ研内で幾人かに訊いてみる。ヤマモト君・マツシマさん・よこやまさんに訊いたり説明したりしてるうちに「それ多重比較じゃん」という突っ込みが入って、教科書やら引っ張り出していろいろと勉強しなおし。統計を真面目にやってない人間にこういう仕事を回すのがそもそもの間違い、と言ってしまうとそれまでなのだけど、それを公言するのは憚られる。ヤマモト君からも「もちょっと基本的なトコからきちんと理解せよ(意訳)」と駄目出しされてしまったことだし、嫌ってないで真面目に勉強する方向で解決を図るべき、だよね。。
夜も遅くなってからウラベさんのところに、このテストは駄目だと思うです、という議論をしに行って、Iδを使ったほうが良いのではという案が採択。ウラベさんとしては最終的には池塘の種多様性を見るようなメタコミュニティを見据えているらしく、…3月頃に下山君に僕が似たようなことを嬉々として喋ったアイデアは既に組み込み済みだったのね。とゆか、芝草プロジェクト自体の落とし所もそういったところなのかなという印象を受けた。
午後過ぎ、研究室。
朝起きてから昨日持ち帰った論文を眺めたのち、本を読んでいたら昼に。
研究計画書を書くために関連文献の収集。結構アンテナが下がっているなあ…。
正午頃、研究室。
池に藻類を放り込む侵入モデル、順番でなくてランダムに放り込むことにして、レプリケートをたくさん採る、ということで。着目点としては一旦固定した競争関係を侵入者が崩せるかどうか、ということになるのか?
作業の合間に雑誌の定期巡回。ピックアップだけして目を通すのは明日以降。
Otsuka (2004), "A theoretical characterization of ecological systems by circular flow of materials", Ecological Complexity(2004), 1:237-252を午後中読む。食物網と物質循環をグラフモデルで記述しているという理解でいいんだろうか。ヘンな論文だなあというのが正直な感想。炭素でほぼ閉鎖循環を考えているので実に違和感があるのだけれど、コンセプト自体はまあ判る。判るんだけども。
この論文は東大の福井さんに教えて頂いたのだが、この雑誌の存在自体知らなかった。他にも幾つか気になる記事が載っている。一応、定期的にチェックするリストに追加しておく。
午後過ぎ、カワタ先生より「またディスカッションが判らなくなった」とのこと、説明。ややこしいデザインと議論でスイマセン。
夕刻前、バイトの登録更新と日時きめ。30分ほど。
メモ:"Cookie Monster crumbles to a healthy diet"
一応、摂餌バランスねた…ということで。単食はイクナイという昨今の風潮に圧されて、彼の有名な歌のフレーズが下記のごとく健康を反映した歌詞になったらしい。
"A Cookie Is a Sometimes Food," where Cookie Monster learns there are "anytime" foods and "sometimes" foods.
彼のアイデンティティというかレゾンデードルとかそういうものは何処に。子供向け教育番組なので、クッキーくらい好きなだけ食わせてやれよという訳にも行かないのかもしれないけど、なんだかなあ。リンク先の記事にも"Sacrilege!"とある。
午後、私用で街へ。
先日よりの引き続いての模試作成。本日で一回目は終了ということで、後日郵送されてきたものに赤入れて返送すれば完了。
先日来より不調だったサウンドボード、いい機会だし買い替え。今回も安物1600円。格段に音質は良くなっていて、しかも5.1ch。我が家に対応スピーカが無いので使われない機能だけども。技術の向上は最底辺でこそ実感できる。
で、今度はキャプチャボードが使えなくなった。全くドリフ仕様(略。というより、諸悪の根源はこいつであるらしい。(数年前だけど)結構高かったのに、毎度こいつのせいでトラブル。素直にカノープスものに戻るかな。同じメーカーのグラフィックボードは好きなんだけど、これも変な仕様が載っかってたりするし。
なんにせよ、新しいのを今すぐ買ってくるのには資金的な律速が掛かったので、お金をためるまでは放置。
朝起き。
暫く買い溜めていた積読本を7冊ほど消化。朝から夜半過ぎまで。半数がラノベだけど、まあそれはそれで良し。
朝起き。午前中、おしごと。
今年度も継続、無理を聞いてもらった形でありがたいことだ。
実習作業中に念仏みたいなのを小声で唱えながら作業しているのがいて(一応授業中ではあるんだけど…)、なんだろうと思ったら、まいやひーを男二人して歌っていた。謎だ。急に暖かくなったからかなあ?
午後、研究室。
終日読書。Andersen(1997)のStoichiometry本とSheffer(1998)のASS本。いずれも一度は通読した筈にも拘らず、ちょっと調べるために要所要所を再読するだけで驚くほどの再発見が! …自分の頭の揮発性の高さを自慢してどうする。幾つか足がかりとして面白げなアイデアに繋がりそうなことが書いてある。
午前中、ガス検査とやらで自宅待機。正午前まで。確かに「まだ」午前中ではあったけども。
正午過ぎ、研究室。
論文直し。作図修正して追記の文章書き。途中でまたあれこれと雑用。途中で気が抜けたんで、昨日のこもごもいう舌も頭も噛みそうな検定のモデルのコーディング。あっという間にできて、あとは分布のマッチについてのDαの設定と実データを貰うだけ。1時間でお釣りどころか10分で済んだ。適当に色々な分布からデータを作って遊んでみる。
そろそろ大阪でインスパイアされたアイデアを幾つかモデルの形にしておきたいところ。ということで、侵入による多様性増加要因のほうのアイデアに関してはすぐに実装できる。流れの中では二種共存問題⇒多種系多様性の低減要因と来て、いかにもベタなんでもうちょい発想にひねりが欲しいところ。
3月頭までやっていたのを上手い事生かせそうな形でASSモデルに組み込めるかも、というアイデア図なんかを早速紙に落書きなどして眺めつつ、さて、どう弄るかな。
午後過ぎ、遅めの昼食を摂っているところに冨樫くん来訪。昨日の今日で早速質問ということで熱心だ。
論文直しつづき。夕刻過ぎにカワタ先生に再送。本日は明日の準備のためにちょっと早めに撤収。
こんどはサウンドカードが唐突に沈黙。デバイスとしては正常に認識され動作しているのに音が出ない。当初はスピーカの不調も疑ったが別音源につなぐと普通に音が出る。あちらを直すとこちらが不調、我が家のPCはドリフのコント仕様か。
本日も長くなった。思考履歴とあるとおり思いつくまま考えるままを整理のために書き連ねているので、考えた分だけ長い。引っ繰り返せば分量が少ないときは考える余裕が無いほど忙しいか、なーんも考えてないかのどちらかだったりする。
午前中、研究室。
論文直し、じりじり。手持ちのは今日中に終わらせるつもりだったのだが。
午後から、研究室のセミナー担当やらその他指針等についての打ち合わせ。毎年、年度頭の恒例。午後過ぎまで。ゴタゴタ揉めて時間が掛かるかと思って午前中に見つけた論文を持っていったが、半分くらいしか読み進まなかった。
午後過ぎ、論文直しつづき。…のつもりが、降って沸いた雑用を片付けてる途中で占部研の富樫君が来訪。以前から頼まれていた下請けモデリングに関するあれこれ。結局彼自身もある程度のプログラミングに関する知識を身に付けたいということで、本日夕刻からはそっち(C++)のレクチャー。超特急でクラスの前までガイダンスして見せて、あとは独習本で見ながらはじめから勉強してもらうという無茶な教え方。
レクチャーの終わりごろになってウラベさんがやってきて、解析方法に関する解説をしてくれた。実データとポアソン過程で得た同数サンプルの分布がマッチするかどうかをKolmogorov-Smirnov(この検定の理屈自体も今更理解したという体たらく)でみて、レプリケートのうち何回はずれが出るかを使って、ある危険率で分布がマッチするかどうかの危険率でもってランダムかどうかの評価をする、というヤヤコシイ検定。
二種類の危険率を混同したのが敗因で、整理して理解できるまでに何回も説明してもらう羽目になって、我が物分りの悪さに閉口する。説明するほうもええ加減にせーよと閉口してたと思う。富樫君はきちんと議論についてきていて、多分僕より先に把握してたような。まあサイワイなことに何とか咀嚼できて、何をどうするかの目処も頭の中で立ったんで、データと細かい式さえ貰えればプログラム自体は一時間もあればお釣りがくるでしょう。
で、とはいうものの、もうちょっとストレートな解析は無いかねえという話をあれこれしつつ、結論の出ぬまま終わったのが夜過ぎ。いやはや。
戻ってきて原稿なおし、手持ち分はほぼ完了。あと幾つか追記するのを考えるのと修正した作図のミスを見つけた箇所の再修正。明日の午前中には終わる。
昨日の話つづき。
栄養塩の持ち込みのケースは多かろうと書いたら、昨日の木庭さんから丁寧なフォローをいただいた(April 5, 2005)。僕がぱっと思いつく程度のことは、既にもっともっと深い所まで追及されているようで。教えて頂いたキーワードで幾つか論文を拾い読み。幾つものシステムのリンクやネットワークまで実際に追跡され始めているらしいというのは、ある面で理論研究側が遅れをとっているなとさえ思えた。
僕程度のアタマでは、それらの「フクザツなモノ」をモデルで表現しようと考えたら(Ecol.ApplとかEcol.Model.とかによく載っているような)「記載的な」モデリング以上を即座には思いつかない。無論そうしたモデルはそれで面白い(個人的な趣味ではハコニワ的あれもこれも全部モデリングは大好きだ)。が、「おらが山」の話でよしというのではガクモンとしての抽象化⇒一般化はしにくい。まあ突き詰めていけばケース・バイ・ケースの側面を生態学は否定出来ないけれど。
システムで見たときの川も面白い。前々から考えていたことではあるけれども、大阪前後の数週間のおかげで、CNRで空間構造を考えるには陸上や河川のほうが遥かにモデル化しやすい印象はより強まった。かまうちさんからいただいたメールにも川でやってみては、という示唆があった。僕が持っている印象としては、モデル対象としての河川系は陸上系と似たような競争(および、その他の)基盤を持ちながら、同時に水系の時間スケールでモノを見れる(かも?)というビジュアルに今のところ一番惹かれる。
というような議論を大阪の自由集会ではもっと掘り下げたかった(+聴かせて貰いたかった)のだけど、…タイムアップになっちゃったのは実に勿体無かったなあ。
私信になってしまうけれど、「テンションは高かったけれど、レスポンスが思った以上に無くてイジケ気味だった」というのが正確な描写でしょうか。幾人かの方にリカバリして貰えたので、現状は高まったモチベーションが残っている好いコンディション。
メモ:"冬季閉鎖道路解禁状況(宮城県)"
お。エコーラインが4.22に夜間以外が解禁、青根道も同日解禁とのこと。笹谷峠は5.10だけど、こっちはブラインドが多すぎてもう行かなくなった。宮城の春は花見桜もいいけれど雪の壁もまた良し。22までに一段落片付けて見に行きたいところ。(雪の壁見るだけなら泉ヶ岳でも見られるけども。)
メモ:"DHMOの国内での使用が全面禁止される見込み"
色々と落ち着いて、今更すらどの4.01ネタを取り上げる。以前化学量論の調べものをしているうちに(例によって横道にそれて)見つけて面白いと思った話。もともとは10年くらい前に当時の中学生が科学賞か何か取った「騙され易さの研究」か何かだった(はず)。普段何気なく見かけるような、環境団体が環境物質に反対する署名を募る文書風なのだが、これを実際に試してみたらほとんどの大人が反対に同意したとか。
公式サイトはこちら。最近は都市伝説やSPAMの一種として語り継がれてるようで、その変遷も面白い。
午前中、研究室。
今日は終日きーきー言っていた(心の中で)。
原稿直しつづき。モデル部完了。コメントに対してもそれなりにもっともらしく反論できたんで、引き分けとまでは行かぬまでも良しとしておく。が。引き続いて昨日返ってきた結果部分、真っ赤。てか赤過ぎ(一番ひどいあたり)。ここまでくると笑うしかないというかネタにするしかなくて、自分で書いた文章が数%しか残ってない(完全文でカウントすると多分0)。一応言い訳だけしておくと、論旨変更はほとんどなく表現に関する修正がほとんど、つまるところ、僕の英作文が駄目すぎという…。あと、競争排除の完了時間については要らんということでばっさりと削除。まあ、その後の議論でほとんど使ってないのだが。…雑誌読みに逃避。
人伝に、木庭さんという方(すぐ下で不勉強と書いてしまった安定同位対比について昔調べたときに拝見したページの方だった)が自由集会の評価をしてくださっている(April 2, 2005付)と聞いて、いそいそと読みに。…何というか面映いです。でも嬉しい。
で、一気にモチベーションは回復し、論文直し再開。我ながら現金な性格である。とりあえず一面赤いところを見続けるのは精神的によろしくないんで、作図に関する指摘から直し。修正された文章のほうにも幾つか気に入らない表現はあって、自分のほうと見比べて推敲。とはいえそういう箇所はあまり無く、修正されたほうが確かに簡潔な表現になっているのを納得せざるを得ないのがほとんどで、実に口惜しい。経験差といわれたけれども、口惜しいものは口惜しい。
メモ:"Introduced Predators Transform Subarctic Islands from Grassland to Tundra"
息抜きがてら(論文直しの逃避)にScienceとかを流し読みしてたのだが…。導入部で、HSS・緑の地球仮説についての言及から始まっていたので、何故いまさらと思いつつ読みはじめたのだが、やられた。実にしてやられた。
まずは概要。アリューシャンの島々では海鳥が鳥糞を落とすことによって陸上植物に対する栄養塩供給に大きな貢献をしている。19世紀後期から20世紀の初頭にかけて毛皮のために狐(=海鳥の上位捕食者になる)が放されたが、幾つかの島では定着が失敗した。狐がいる島では植生が劇的に変化した(草原⇒ツンドラ)が、それらの島で施肥すると狐のいない島とよく似た植生になった。この理由は従来のHSSでの捕食による植食圧の抑制ではなく、捕食によって海鳥からの栄養塩供給が抑制されたことによって説明される。
何がしてやられたかというと、植食者による栄養塩再供給が上位捕食者によってコントロールされるというモデルを考えていた。のだが、ほぼ同じことを実際のフィールドでやられてしまった(この論文は再供給でなく外部からの持ち込みだが)。捕食者−植食者−植物というtop-down cascadeな直列の食物系ではなくて、捕食者−被食者=栄養塩供給者⇒植物という、面白い系を見つけてきているのも大きい。陸上生態系での鳥糞による栄養塩供給が面白そうだと思った矢先に読んだこともあって、そっちからもダブルパンチ。
確かにねぐらとか考えると、生態系における動物による栄養塩供給は閉鎖系内での循環だけでなく、こうした持ち込みというケースもかなり多いはず。というか家畜と牧草地の研究で空間的な効果も込みで幾つか見た記憶がある。この島では糞落はスポットにならないみたいなので異質性に絡めるのはやってないようだけど、それは僕のアイデアのほうの話で、この話には全く関係ない。
もう一点、栄養塩が何から由来するかについて安定同位体から解析していて(多分)、結構な行数がその説明に割かれている。セミナーとかで説明された以上の体系的な理解をしていないので、このへん出てくると自分で理解できる内容が途端にアヤシクなる。大阪で聞いた話でも何度か出てきたなあ…。ストーリーは把握できたとして、データ面からの評価ができませんではマズイよね。
おそらく今後のCNR研究で、実際に野外の定量的なデータ等を知りたいと考えるようになったら避けて通れない技術だろう、というのもある。
正午前起き⇒研究室。
春眠何とやら。
ちと思いついて必要になったコーディング。ポアソン分布を自動的にデータ作成⇒作画するプログラム。前に作った乱数生成関数my_rand(uboud, lbound)はなかなかに汎用性に長けたスグレモノで、大したことをしない割にあちこちで重宝する。で、今回もこれを使って10分ほどで作成。1万回レプリケートをとって平均頻度分布に分散を乗っけてみると、分散がえらくでかい。計算は間違ってないはずだし、試行回数を変えても頻度分布は変わってないんだけど、こんなものか?
…とか書いてたのだが、期待値と分散が等しくなるのを失念していた。不思議でもなんでもないというオチが待っていたのだった。
午後過ぎから論文直し。論旨は固まっているのでほとんど動かないが、英作文の直しが多くてへこむ。が、確かに納得できるものも多いので勉強させてもらっている。論旨そのものについての言及や追加せよという指示については対処したり反論を準備したり。そうこうしているうちに、カワタ先生からの追加が到着。早くこなさないと手が廻らなくなりそうだ。
生態学会のポスターおよび自由集会についてのレスポンスをいただいて非常に嬉しい。特に自由集会については討論が省略されてしまったこともあって、話したら話したなりに(駄目出しでも良いから)リアクションが貰えないのは寂しい。
朝起き。
昨日よりの作業つづき、の合間に部屋の掃除をしたり買い物に行ったり。
午後過ぎより友人が遊びに来て夜過ぎまで。その合間にOSの入れ替えやら必要なソフトのインストールやら作業を続けて、夜半過ぎには大方完了。結局、HDD(80G)⇒HDD(250G)の交換になったんで実質は170Gほどの増設、のはずだが既に空き容量が100Gくらいまでに逼迫している。何故だ。
朝起き。午前中、研究室。
冬も多分あけたことだしバイクを洗車。結構汚れていたのに加えて、消耗品の類がタイヤ以外軒並みヤバゲ。
正午過ぎ論文直しつづき。これだけ真っ赤になってると正直なところウンザリというのはある。
午後過ぎ、私用で駅裏。模試作成のつづき。夕刻過ぎ終了。
部品屋でHDD(250G)購入、15000円。自宅のPCのHDDの空き容量がとうとう200MBとかいうかなり差し迫った状況になってしまい、仕方なしでの出費。
自宅に戻って増設作業。とくに問題なし…かと思いきや、128G以上の容量を認識してくれない。BIOSをアップデートしても(COMSのSUMが合わなくて起動しなくなったりとかしながら)認識してくれず。どうやら調べてみるとFAQであったようで、Windows2000で大容量のHDDを利用するときには一部のレジストリを書き換える必要があるらしい。
夜半からデータの移動作業。いっぱいありすぎて延々と掛かる。
今日から新年度、4月。課程の最後の年。
大阪の生態学会で使った資料を公開。
朝起き。午後、研究室。
大阪から送った荷物の中にPCも入れていたので、午後過ぎに届くまで色々と作業できず。大阪に行っていた間に溜まった雑用片付け。ほぼ完了。
{kind=link}
{kind=link}