99�@�_�����e�����FCNR�ɂ�鋣���r���x�����f��
85�@���f���F"The specialization v.s. generalization in dietary stoichiometry"(����)
60�@���f���˘_���F��ԃp�^�[���������f���̂܂Ƃ�
50�@���f���F�ő������蒆�����f���i�g�K�V�����j
20�@���f���F"The specialization(��)�ˊg���F��ԒT��
15�@���f���F"The specialization(��)�ˊg���F�H���ԍ\��
10�@���f���F�~�W���R�̐����K���ƔɐB�����x�̊W
-5�@���f���F��ԉ�݂Q�틣����CNR���f�����ˁiASS or PoE����̃A�v���[�`�H�j
�@�ߌ�߂��A�������B
�@�I���A������������낤��B�s����ȕ���ʼnE�������悭����Ȃ��̂ƁA�����O�̌����\�͂̒Ⴓ�����ւ��āA�v���悤�ɕ������W�܂�Ȃ��B
�@�[���A�֓lj�B��T�̑����Ŗ�艉�K�B�s��̃x�N�g���v�Z�ɑ���C���[�W��������ƕς�������Ƃ������āA�X�e�[�W�\������荞���f���Ȃ�Ă̂��ʔ�����������Ȃ��A�ȂǂƎv���A�ꎞ�ԂقǁB
�@���A�������B
�@�ߑO���A�Z�~�i�[�B�S���͖l�ŁA�挎���炢����V�����l���Ă��郂�f���̘b�i�g���������j�B���̃A�C�f�A�ł�����o���邱����o����Ƃ����Ƃ���ɊႪ�s�������āA�̐S�̑������܂���������ɂȂ��Ă���̂��Ղ���Ă��܂����B���ǁA�x�[�X�R���Z�v�g��������Ƌl�߂Ă���A�b�͂��̏�Ői�߂�ׂ��Ƃ����ᔻ�ɏW�����闬��ɂȂ��Ă��܂����B���\�̂��ƂŁu�r������́A�����v�悶��Ȃ��Ėϑz����ˁv�Ƃ������]���������������A�S�����̂Ƃ���B���ȁB
�@�Ƃ�����ŁA���ȗ��A�R�����Ԃ�̃v���[���e�[�V�����̎��ȍ̓_�B�����̍\���̕s�����i�|�P�O�j�A�v���[���e�[�V�����ɂ�������p�̓K���i�|�P�T�j�ƒ��J���i�|�P�O�j�A�����R���Z�v�g�̐����i�|�R�O�j�A������̃��A�N�V�����i�{�P�O�j�Ɨ���x�i�|�T�j�B�v�T�O�_�B������̃��A�N�V���������\���������A�̓_���������Ȃ��Ă�悤�ȋC�����Ȃ��������B
�@�A�C�f�A�͈����Ȃ��Ƃ����]����������������ŁA�����ʔ������T�b�p�����Ɖ]���Ȃ����������͋~�����ȁB�܂Ƃߕ�����������ƍl����ΐ����͂̂���ʔ������f���ɏo����͂����B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@�Ƃ����킯�ŁA��ԍ\���Ƃ��w��x�ɑ���O�Ɋ�{�R���Z�v�g���ł߂�ƁA��������B���h����Ă��܂����B
�@����Ńh�R�ɖ�肠�邩�Ƃ����Ƒ傫���͓�B�B�{�����ɂ�邱�̕���ւ̃C�m�x�[�V�������Ȃ�ł��邩�Ƃ����Ƃ���m�ɒł��Ă��Ȃ��̂��ЂƂB�@���ĂȂ����Ȃ疳���ɂ���A�Ƃ����z���B����͖l�̕��s���ɋN������B�����ЂƂ̓R���Z�v�g�ɉ������\����������Ɨ��Ă��Ă��Ȃ����ƁB����Ă݂Ȃ��Ɣ���Ȃ������̌��������邾�낤���A����͂������������Ŗ����Ǝv������A���͖l�������̑S�̑����\���ɔc���ł��Ă��Ȃ����߂��낤�B����͖l�̍l�@������Ă��Ȃ��؋����i�Ƃ������́A��͂�Ⴊ��Ɍ��������ĂāA�n�ɑ������ĂȂ������Ƃ������Ƃ��j�B
�@�J���^�搶�Ƃ̒��ڏ����͎b���C���^�[�o����u���Ă���B�O���킽�鍡��̓{�������ɋ߂������킯�����A�c�������́B
�@�ߌォ��C�������Ă����B�g���̂Ẵv���O���������Ă݂���B�����߂��B
cout<<"STDOUT"; cerr<<"STDERR";
���������Ƃ��āA(bash/zsh�ł́j./a.out 1> 1.txt 2> 2.txt�Ǝ��s�����1.txt�ɕW���o�͓��e���A2.txt�ɕW���G���[�o�͓��e�����ꂼ�ꃊ�_�C���N�g�����B*csh�n�͐��䂪��₱���������L���͂��邪�A��肩���������ĂȂ��B
�@�{���̗p�r����͊O��Ă�悤�ȋC�����邪�A������g���ƌv�Z���ʂ̏o�͐�̐U�蕪�����i�Q�܂ł́j�蔲���ł���B
�@���߂���A�������B
�@�ۈ���A���\�����̍쐬�ƏC���B�[��������R�����������̂Łi���삳�Ăт����Ă��ꂽ�A���Ӂj�A����ɑΉ����Ă���ɏC���B����͖l�̕��s���ɂ���ĕs�\���ȑΏ��Ȃ̂����A���ɔ��\�ɂ͊Ԃɍ���Ȃ��B
�����������_�ŁA���f���Ɋւ���l�@�̋l�߂̊Â����F�X�ƘI�悵�Ă���B������x�A�C�f�A�̕��������o�����̎��_�Ől����ӌ���Ⴆ��̂͂��傤�Ǘǂ��^�C�~���O�Ȃ̂����B�I����Ă݂���S�s�ł����ƂȂ�Ȃ��悤�ɂ�����Ɣ��\���Ȃ��ƁB
�@�ߑO���A�������B
�@�o�C�N���ɂ���ĕ��i�����̏I�����SV�������グ�Ă��猤�����B�����Ă�������A�r������J�ɍ~���ĔG��l�B���ǁA����̃G���W���I�C�������ɉ����āAF�u���[�L�p�b�h�����AR(�u���[�L�p�b�h�A�p�b�h�s��*2�A�u���[�L�L�����p�[�̃I�C���V�[���A�u���[�L�t���[�h)�����B�L�����p�[���̂��K�тĂ��̂��Ǝv���Ă����ǃs���������̂��B�S�����݂��݂�25,000�~�ȓ��Ɏ��܂�������A�܂��悵�Ƃ���B�ł����i��͂��ƈ������ǁA�H����������˂��c�B
�@���\�����������蒼�����Ă���v���O�������������B
�@Denno and Fagan(2003)��ǂ�ł��āA���i������H�ׂĂ�A�������f���ɂȂ�ƐA���i��q��ԕ��j��H�ׂ�Ƃ����L�q���ǂ��ɂ��D�ɗ����Ȃ��āi���ɉԕ�������قǍ��^���p�N�Ȃ̂��A�Ƃ����_�ɂ��āj���R����ɐu���Ă݂��Ƃ���A���͎�q�����łȂ��ԕ������Ȃ�CN�䂪�����a�ł���炵���B�����Ă��������������ɂ͉ԕ��̉h�{�����͏Ă��T���}�Ɠ����x�i���������b:�^���p�N��j�Ƃ���B�Ă��T���}�Ƃ����\���͂�����Ɩʔ����i���Ԃ�h�{�w�̖{������������Ă����f�[�^�Ǝv����j�B�܁A�e�Ɋp�A�ԕ��̉a�Ƃ��Ă̎��ɂ��āA�Y���������܂߂Ă����Ȃ蒂�f�䂪�������Ƃ�m��A�Ȃ�قǔ[���ł����B
�@�ߌ�߂��A�Z�~�i�[�i�}�j�B
�@��T�ƃ����o�[�\���͕ς��Ȃ��̂����A���T�̓Z�~�i�[������c�B
�@�O���A��t���̖���S���B
�uShell choice in Paguruslongicarpus hermit crabs:does predation threat influence shell selection behavior?�v�i�o�T�s���j�̘_���Љ�B�����̃��h�J���Ŏ����悤�Ȍ������l���Ă���Ƃ̂��ƁB�B�z�����h�J���͎���������k��I�ԍۂɁA�i�Ȃ�ׂ���ɂ����ߐH����ɂ����j���̖������̂�I�Ԃ��߂Ɋk�ׂ邻�����B���̕ߐH�҂ł���~�h���K�j�i���������ȁH�j�̋��饂��Ȃ��ɂ���āA�k�ׂ鎞�Ԃ��ς��Ƃ������e�B
�@�D�ɗ����Ȃ��̂́A���̏�Ԃɂ�������炸�A�ߐH�҂�����e���ɂ���ĉ�����鎞�Ԃ���������Ƃ������ʂ��o�Ă���_�ŁA�����I�ɂ͕ߐH�҂�����Ȃ牽�ł��ǂ�����Ƃɂ������ł��Ȃ��悤�ɂ���悤�ȋC�������ŁA���̌��ʂ͎ߑR�Ƃ����A����ɑ���l�@���u�k�̈��S���ɑ��郊�X�N�]���ɂ�蒷�����Ԃ��|���邩���H�v�Ƃ����A�Ȃ��悭����Ȃ����̂������B
�@�ŁA������C�ɓ���Ȃ��̂́A���̌����ɂ����Ă��A�w���݂��邫��[�x�����ɐ��ސ����ɂ͕��ՓI�ɓ����Ƃ������ᔻ�ȑO��͂��܂��Ă���_�B���̍���䂦�ɁA��̓I�ȕ��������肳��Ă����͂قƂ�ǖ����i��������H�j�B���̌����ł���ɘR�ꂸ�A�؋�����ߐH�҂̋����c�萅���g������s�����ς�����A����́w���݂��邫��[�x��F�����Ă���ɈႢ�Ȃ��Ƃ������W�b�N�ȖB
�@�Ȃɂ�����������Ƃ�������͖����āA�Ⴂ���o�Ă镪�ɂ́u����낤�ˁv�Ƃ����[���͂���B����͂����̂����A�ł̓f�[�^�ɍ��������Ȃ������ꍇ�ɁA���ꂪ�w���݂��邫��[�x���̂��̂����������̂��A����Ƃ����邯�nj����Ȃ������̂���ʂł��Ȃ��͖̂�肶��Ȃ��낤���B�ߐH�҂����Ă����Ȃ��Ă��s�����ς��Ȃ��A�Ƃ����_�݂̂ɂ��Ă͊m���ɖ��͖�����������Ȃ����A�ߐH�҂��i���Ȃ��Ƃ����w�I�ɂ́j�F�����Ė����̂ƁA�F�����Ă邯�Ǎs����ς��Ȃ��A�Ƃ����̂ł͂��炢�Ⴂ������Ǝv���B
�@�ƁA�����܂ōl���Ă������A�w�Ȃ��Ⴂ���o�Ȃ������̂��H�x�Ƃ����l�@�����Ă錤�����́A�����ɂ́A�w���݂��邫��[�x���g���ĕߐH�҂̑��݂��r������s���ɍ����o���A�Ƃ��������̕��������B�o�Ȃ��������̂Ɋւ��ẮA�i�Ȃɂ���������܂���ł����Ƃ����_���ɂ͂Ȃ�ɂ�������j���������f�[�^�������Ă���\�������邪�A�f�[�^�ɈႢ���o�Ă���Ȃ�A��q�̂悤�ȐS�z�͞X�J�Ȃ낤���B���̎�̌�����ǂ蕷�����肷�邽�тɂ������Ǝv���̂����A�z���g�ɖ��Ȃ��̂��ǂ��������ɐ����ł��Ȃ��܂܂ŋ���B
�@���Ɂw���݂��邫��[�x������̂͊m���炵���Ƃ����̂��x������Ă��鐶���͕ʂƂ��āA�͂Ȃ���w���݂��邫��[�x�̑��݂�O��Ƃ��Ă��錤����@�̓C�}�C�`�D���ɂȂ�Ȃ��B�\�������Ŋm���߂Ă��錤���Ȃ�s���͊����a�炮����ǁA�w���݂���Ƃ����Ă���i�؋������Ƒ����Ă���j���A���̐��͕̂s���x�Ƃ����̂��C�ɓ���Ȃ��Ƃ��������ȋC������B�����Ƃ��A�ł͑���ɂ�����g���ׂ��A�Ƃ������������Ă�̂ł��Ȃ���ŁA����I�Ɍ������Ƃ����Ă�ȏ�́A���ݓI�Ȕᔻ�Ƃ͂����܂��c�B
�@�㔼�A��t���̎O�Y�N���S���B
�����̥�̌Q��R�~���j�e�B�ɗ^����C���p�N�g�ɂ��Ẵ��r���[�Z�~�i�[�B�ʔ������������ɂ��Ȃ������A�m���������Ă���^�C�v�̃Z�~�i�[�Ȃ�ŁA����͂����ɂ͂������ǂ��܂藍�߂Ȃ������B
�@�߂��Ă��ăR�[�f�B���O�����B
�@���낻��^�C�����~�b�g�������Ă����킯�ŁA���o�C�ȁ[�B
�@�c�Ƃ�����Ŗ�߂��܂Ŋ|�����āA�ЂƂ܂��`�͏o�����B���j�b�`���Ɖa�l�������̃g���[�h�I�t�̕�����ӎO�p�`�ŋL�q����̂͊Ԃɍ��������������B�ŁA�K���ȏ����ł͒m�点�Ă݂�ƁA�\���ǂ��蕝�͏��X�ɋ��܂���A�ʒu����܂炸�t���t���s�����藈���肵�Ă���B�����͂悭����Ȃ����A�����炭�͈�ԍD�݂̒����l�ƍ��E���[�ł̌����̒l���i�^�������`�ɂ��Ă���̂Łj���������߂��낤�B���̕����͐^����ɉ��P���Ȃ��ƃC�J���ȁB
�v���������v�Z���Ԃ��|���肻���Ȃ�ŁA�Ƃ肠�����v�Z���d�|���Ė�߂��ɓP���B
�@�����N���B
�@�N�����͗ǂ����A�W�F�l�V�X�͖{�����S���t�̂��ߕ������Ȃ������c�B2�T�Ԃ�̃q�r�L��������\���Ă��猤�����B���̊قȓW�J�͒��Ղŗ\�z�ł������ǔR����B���T���y���݂��B
�@�ŁA�����Đt�R�B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@�ߌ�߂��ɂ͎������o�����B���ăv���O�����̑����������Ƃ��悤�B
�@����trade-off between handling efficiency and niche width�͓ӎO�p�`�ŋL�q���邱�Ƃɂ����B��܂ł���ĕ����������̂œP���A�����ƂɃX�C�b�`�B�c�̂��肾�����̂����A�������ł̗[�т��������B��тɊ��̃^�^�L�A���ߎI�B��y���ɂȂ�܂����B
�@�����ƁB
�@�����̂����h�C�̂Ŏ���ŃR�[�f�B���O�����B��T�ԑO�ɏ������v���O�����̒��g�Ȃ��R�Y��Ă�̂ŁA�����܂ł���Ăč����牽�����̂��R�����g�����Ȃ���v���o���̂ɂ��炭������B
�@�K���x�̌v�Z�������̑������A��܂ŁB
�@�ߑO���A�������ƁB�ߌ�A�������B
�@�I����Ă���o�C�N���B�L�����p�[�̌����p�������Ă�����ŗa����B���j�Ɋ����̗\��B���̂܂ܓk���ɂĐt�R�܂ŁB�U���Ƃ����̂ɂ����Ԃ�Ə����B����Ȃ��̂������낤���B
�@�ߌ�A�������B
�@����̑����A�R�[�f�B���O���ǂ�ǂ���ɂ��ꍞ�ށA����������܂����B
�@�ߌ�߂�����_���[�~�i��j�B�{���̓}�L�m����ɂ��ALee and Frost(2002), "Morphological stasis in the Eurytemora affinis species complex (Copepoda: Temoridae)", Hydrobiologia(480):111-128.�̏Љ�B�z���g�̓E���x����Ɠ�l�̗\�肾���������ԓI�ɖ����Ƃ������ƂŁA���͐��j���ɑ���ɖl���o���Ȃ����Ƃ����̂�����Ă��Ă����B���A�������ɗ��T�̏������l����Ɩ����������̂Œf������������ׂŁA�{���̓}�L�m�����l�B
�@���e�Ƃ��ẮA�P���~�W���R���L�͈͂ɃT���v�����O���Ē��ׂĂ݂�ƁA�`�Ԍ`���̂������x�ɔ�ׂĈ�`�I�Ȑi�����x�̂ق������������Ƃ������e�B�ŁA���̌����ɂ����Ă͉��Ō`�Ԍ`���̕����ۑ����ꂪ���ɂȂ��Ă���̂��H�Ƃ����c�_���������A���������B
�@���I����Ă݂Ă̊��z�B���A���^�C���ɏ������āA���A�܂Ƃ��ȋc�_������ɂ͂��Ȃ荂�����x���i�Ƃ������W���͂��j��v������Ă邩���B�茳�ɗ^�����Ă���̂��_�����̐}�\�����Ȃ̂ŁA���\�҂ɂ������ɕK���ɐH�����Ă����Ȃ��Ƃ�������u���Ă������B�������s���ȓ_����������C���^���v�g���Ė₢�������悢�̂����A�r���ōl�����e�Ɉ�ꂽ�r�[�Ƀq�h�C�ڂɂ������BM1�Ƃ�B4�̊w���̐l��ϋɓI�ɋc�_�Ɉ������荞��ł������߂̂͌��\��ς����B
�@�[���AJeconet�ɂē���̓��c��������T���Ă�����B���ׂ��瓌�k��̐��w�������̑����Ƃ킩�����̂ŘA�������グ�āA���ʂ��ėX������B�߂��Ă��č�Ƃ̑����A��܂ŁB�v�|�������Ă�����t���ς�鍠�P���B�茳�Ɍ��ʂ��o�ĂȂ��̂łȂ�Ƃ����g�̂Ȃ��X�J�X�J�ȗv�|�Ȃ̂���Ȃ��B
�@�ߑO���A�������B
�@���������|���Ĕ��\�������B��g�Ƃ��Ă͂ł��Č�͔����߂��葫������B�܂��A�܂����X�e�b�v�̌��ʂǂ��납�v���O�������r���܂ł����o���ĂȂ��Ƃ����w�傫�Ȍ��x�����܂��ĂȂ��킯�����c�B
�@�[���A�J���^�搶�Ƀ��f���̑�g�̑�g�ɂ��ď����b���Ă��炤�B��Ԃ���̃��f���̑O��ŐF�X�ƍ��肻���ȉӏ���������A�����A�h�o�C�X���Ⴆ�����Ȋ����B���̂��߂ɂ��A���̃��f����l���ʔ����Ǝv���Ă��邱�Ƃ�������Ɠ`����̂��̗v���B
�@�[���A�֓lj�B�I����Ă���܂��b����������ƎG�k�B����ɂ킽���Đ[�������Ă�����Ȃ̂Řb�����Ă��ĕ��ɂȂ邱�Ƃ������B�߂��Ă��đ����A��߂��܂ŁB
�@���f�����̂̉��P�ĂɊւ��Ă��������J���^�搶�Ƃ̉�b�Ŋ��Ɍ�����������������A������ł͂������ɊԂɍ���Ȃ��B�Ƃ肠�����͌��ʂ��̂��߂ɑ��i�K�܂Łw���g�݁x�Ō��ʂ��o���Ƃ��悤�B�������ɂ͗v�|���o���˂Ȃ�ʂ��A���ʂ͏T���ɂ˂����ނ����Ȃ����B
�@�ߑO���A�������˃Z�~�i�[�B
�@�{���̓��U������̒S���Ō����Љ�B�^�ߍ��̃R�E�����̈�`�I�\����(1)�n�r�^�b�g�i�A�����H�j�̒n���I�ȍ\����(2)�̌Q�T�C�Y�̌����̗��j�I�v���̗����f�������ʂɂȂ��Ă���Ƃ������e�B�b��̓��e�ɐe�����Ȃ��̂Ɖp��͂̒Ⴓ�����ւ��āA�p�����[�^����̂��߂ׂ̍��������ɂ��Ă����Ȃ������̂ƁA������Ƃ������₪�o���Ȃ������B���낢��ƏC�s������Ȃ��c�B
�@�ߌォ���܂Ŋ|���Ĕ��\���e�̍\�����B��܂��ɂ͏o�����A���A���\�{�����[��������B����͓��ɍ��܂Ō����ɂ͈�x���b���Ė����V�������e�ł��邵�A�l�Ƃ��Ă͔��ɖʔ����Ǝv���Ă���b�Ȃ̂ŁA���ꂪ�����Ɠ`����Ėʔ����Ǝv���ĖႦ��Z�~�i�[�ɂ���ׂ��A�Ȃ邾�����J�ɍ\�����߂Ă������Ƃɂ���B
�@�����ɍŒ���K�v�ȃ��f���̃A�E�g���C���ƌ��ʂ��̂��߂̌��ʁi�Ȃɂ�������悢���j���ϋl�܂��Ă����B
�@���ɕ{���ʂ���A���Ȏw�������ł����̂Ŗ{���͖{�����J���Ă邤���ɒ��֊��ׂ��A���t�̕ς��ʂ����ɓP���B
�@�����
�@�ߑO���A�������B�ߌ�o�C�g�B
�@�v���O���������̍��Ԃɗ��T�̔��\�Ɍ����ăX�g�[���[�\�����l�����ɉӏ������B�O��܂ł̘b�i���ނ̑��l����CNR�Ƌ�ԍ\�����^����e���̓z�j����A������Ă�b�ɂǂ��q����̂��Ƃ����Ƃ���܂łЂ�����߂�ƌ��\�ȕ��ʂ̃C���g���_�N�V������p�ӂ��Ȃ��Ƃ��߂����B���\��邱�Ƒ����c�B�ߌ�߂��o�C�g�B
�@�[�����A�ߓ�����ɂ��Z�~�i�[�B
�@�J���^�搶�Ƃ̋��������̑ł����킹�Ő��ɂ���ꂽ�ߓ������ɂ��u�_��ȐH���ԁv�����Ƃ���ɂ܂��ŋ߂̋c�_�ɂ��Ă̏Љ�B�O���͂���܂łɂ����������Ƃ̂�����e���������A�㔼�͍ŐV�̎�������������ɔ���Ղ��v���[���e�[�V�����������B�H���Ԃ̍\�z�Ɋւ���b�ł́A���l������Ă��郂�f���ƃR���Z�v�g�̒ʂ���Ƃ��낪���\�����ė�⊾���̂������B
�@�Z�~�i�[�̒��ō������ɓǂ�Williams and Martinez(2000), Nature(404):180-183�ɂ��ċߓ������x�����y����Ă��āA�l�̗������܂��܂��s�\���Ȃ��̘_���̘b�́A����̕����������Ă��������Ŏv�����ȏ�ɂ��Ȃ�d�v�ł���ƍĔF���������B�ߓ�����̞H���A���͂��̘_���ɍڂ��Ă������ł͊��S�Ȓǎ����o���Ȃ�����A���ڃR���^�N�g���Ƃ��Ċm�F����Ƃ̂��Ƃ������B
�@�̂��A�ő��֔��܂�Œ����ɍs���Ă����E���x������������Ē��ŐH���A���^���̐H�ׂ���ē����i���O�����O�j�B�I����Ă���A�ߓ�������h�܂ňē�����܂ŁA�������Ă獡����Ă郂�f���ɂ��ď����b���Ă��������B�������̃O���[�v�������悤�Ȃ��Ƃ��l���Ă��邻���ŁA�ł��邾���}���������ǂ��Ƃ����A�h�o�C�X�������������B
�@�������ɂ͖߂炸�A���̂܂���֓P���B
�@�ߌ�A�������B
�@���N���Ė{��ǂ�ł��炵�炭�����ƁB�ڏ��������i�K�Ō������A��Ǝc���Еt���B�[��߂��܂Ŋ|�����ăX�N���v�g�̋@�\���������B����͂�A���ǁA�T���ƍ����ۈ�����₵�Ă��܂����B
�@����̋��P�́A�^�R�Ȑv�Ǝ��������Ă��܂��Ƃ��Ƃ��玩���̑�������������A�Ƃ����̂ƁA�����̑z�������������郆�[�U�͂������邩��A�e�X�g�͏o�������Ȗ��ɂ���āA���A�����e���₷�����̂�p�ӂ��邱�ƁB�܂��A�]�������Ȃ�ȒP�����A�Ȃ��Ȃ��B
�@�Z�~�i�[(�})�́A�E���x�������t��������炸�A�����ɏo�����Ă�q�g�����������̂Ŏ��T�ɉ����B�Y��Ă��킯�ł͂Ȃ��̂����ǁA���傤�ǃX�N���v�g�̋@�\���������Ă���ŏ��������B�ŁA���T����t����蕔����Ƃ��ɕs�݂������C��������c�ǂ�����낤���B
�@�[������ǂ����T�[�o�̒��q�����������A���炭���Ԃ��o�߂���ƃT�[�o��������B���ɕ��ׂ��|�����Ă���l�q�������A�����̃e�X�g��Ƃ������Ƃ��v��������X���Ă����̂����A�b�����Č����������B�}�T���N���T�[�o�����Ă����ăS�\�S�\�ƍ�Ƃ����Ă����̂����i��[�������Ă�����ɗ������j�A���ܓd���^�b�v���R����Ă��̓s�x�T�[�o�𗎂Ƃ��Ă����炵���B���킭�w���������Ύ��X�ȂJ�b�`���Ɠd�����悤�ȉ������Ă܂����x�Ƃ̂��ƁB���c���A���@�́@�@���@���@��@�[�B
�@�܂��A�ڐG�ɂ��s��������d���ɂȂ��ł����l�ɂ��ӔC�͂���̂����B�����B���������Ɍ�������Ƃ�����B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@�{���͊ۂ������t�����B���낻�딭�\�����̕��ɂ����|����Ȃ��Ƃ܂������ƂɂȂ�c�B�ЂƂ܂��́A�܂����f���̂ق��ɏW���ł������ł悩�����ǂ������B
�@���\�\�����l���A�T�Ȃ��܂łɃ��f�����o����g�R�܂őg��ŁA���\���Ė�������Ƃʼn��߂ăJ���^�搶�Ɛ퓬���邱�Ƃ�z�肵�āA�T�Ȃ�����T���ɂ����Ę_���\�����܂߂����\�����A�Ƃ����蔤�ł����Ƃ��悤�B
�@�����ƁB
�@���ł̓q�r�L����݂̂Ȃ炸�W�F�l�V�X�������ǂȂ̂ŁA�{���͗����Ƃ������Ȃ��B�G�E���J�����ςĂ���܂��s��Q�B�T���߂̊��͌����[�B
�@��k�͂��Ă����A����̑����B�@�\��ʂ���Ƃ������ƂŃ`�g�d�l�ύX���l���Ă݂�B
���́ˏd���o�^�̊m�F�˓o�^�ƕۑ��˃��[�����M
���o�^�ҔF�˓o�^���e�ǂݍ��݁˕ύX/�폜�˃��[�����M
���o�^�ҔF�˓o�^���e�ǂݍ��݁˕\��
�Ƃ����킯�ŁA���͐V�K�ɏ������������͂����킸���Ɍ�����A�����͒P��̃X�N���v�g�ɋl�ߍ��ލۂɁA�ׂ̃��W���[���̈ꕔ���p���Ȃɂ�炵�ăp�b�L���O������������ׂŁA���X�X�p�Q�b�e�B�ȍ\���ɂȂ��Ă���B�ߌ㒆�A�X�N���v�g�̏��������B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@�悭�悭�l���i�߂�ƁA�j�b�`�ւ̓����z���͓ӎO�p�`�ł���K�v����Ȃ��A���ƍ������ʐψ��Ō��܂钷���`�ł悢�B�ނ��낱�̕������spcialist/generalist�I�ȓ�����ǂ�����킵�Ă���igeneralist�͂ǂ̎����ɂ������������������ɂЂƂ�����ւ̓����ʂ͏��Ȃ��j�B
�@�A�C�f�A�̓������Ă������A�������A��͓�{�����Ȃ��̂Ńv���O�����������ɖ����B�����ɂ͎����̃Z�~�i�[�S��������Ă��邪�W���n�Ȋ����B����͂�B
�@�����A�Óc�N���̃��[�����m�F�B�t��cgi�Ɋւ��āi���[���A�h���X��ύX���悤�Ƃ��Ă��o���Ȃ��j�s��̎w�E�B�����m�F���Ă݂���m���Ƀ}�Y�C�B���A�����炩��̐����s���̂܂܃A�i�E���X���o���̂��X�Ȃ�}�C�i�X�v���ɓ����āA�͂���ɍ������łǂ������Ă���B
�@�ߌ�߂��A�������B
�@�����C���̂��߂ɂ������e�X�g������A�Óc�N���o�����G���[�i�X�V�o���Ȃ��͂��Ȃ̂ɕБ��̃f�[�^�����X�V���ꂽ�B�j���̂��͍̂Č��ł����B���̖{�l�ɂ��Č��ł����B�䂾�B���[���A�h���X����̐v�Ɋւ��ĂȂ̂����A�G���o�O�ł͂Ȃ��Ė��炩�ɐv�~�X�B�v�i�K�ŋC���t���Ȃ������̂͂����ɂ��Ԕ��������A���̂ւl�̔��z�̌��E�Ȃ낤���B
�@�ŁA�܂������_�ŊY������o�^�҂͓�l�i����l�����₢���킹�̃��[�������ɂ��Ă��āA������l�͉����ς݁j�Ȃ�ŁA�u���܂̂Ƃ���́v����܂���Q�炵�����Q���Ȃ����A���̐�A�o�^�Ґl����������ɂ�ăg���u�����������邾�낤�Ƃ������Ƃ�����Acgi�X�N���v�g���@�\���Ƃɂ������ɕʂ��邱�Ƃʼn������悤�Ƃ������ƂɂȂ����B���\�啝�Ȑv�ύX�Ȃ�ő�ς��B
�@�ŁA��o�^�t�H�[���œ��͓��e�ɃG���[������̂ŏC�����Ă��������v�Ƃ���ꂽ�Ɠ{���Ă����郁�[�������������B���e�ɃG���[���o���Ƃ��������ĂȂ��̂ŁA�����̒N���ǂ����ė~�����̂�����Ȃ��B���[���ڎ�����Óc�N�����f���Ă���̂ŁA�u���傪�������������������悤������ӂ��č����グ��v�Ǝw���B
�@�����Ƃ��Óc�N�͗ǂ��o�����l�Ȃ̂ŁA���̎w�������̂܂܉L�ۂ݂ɂ�����͂����A������Ɖ����̒N���ǂ����ė~�����̂���₢�Ԃ����e�̕ԐM�������������B�ǂ������̐��ɂ͈��̊����ŁA�V�X�e���S���͋@�B�������������Ă���Ǝv���Ă��������������悤�ŁB�����Ƃ��@�B�͋C�𗘂����Ƃ������Ƃ����Ă���Ȃ�����A�Ȃ�����t�H�[�}�b�g�ɑ���Ȃ��Ɠ����Ȃ��͂��Ȃ��ǂ��B
�@�v�͢������l�Ȃ���A�m��Ȃ�����ւ̎莆�̕��ʂ��炢������Ə����Ȃ����棂Ƃ����b�B�q������Ȃ�����u�ł��Ȃ���ł����ǁ[�v�����̃��[���͒p���������B�����̓{���`���ė������ė~������A����ɕs���Ȏv���������Ĕ�������������̂ł͂Ȃ��A�Ȃ�ׂ����k���Ă�����Ĕ��ȂƉ��P�𑣂��ׂ��ŁA���̂��߂ɂ͂Ȃ�����Ɏ����ȕ��ʂ��K�v�ɂȂ�B���̂��Ƃ͍����ɏ����Ă��镶�͂ɂ��Y������B�Ȃ��J�������Ď����𐳓�������悤�ȓ��e�ɂȂ��Ă��܂����A�ƁA���������߂c�B
�@�{���͎��p�ŗ[�����߂ɓP���B
�@���N���B�ߑO���A�������ƁB�ߌ�A�������B
�@�{���͗֓lj�i��j�͐V�K�Ɍ����ẴC���^�[�o���B�ŁA�ߌ�߂��Ɍ����A�ꎞ�Ԏ�ōċN���B���ɐ������Ԃ͌����ĂȂ����ǁi���炵�悤���������j��T�Ԃɐ��������ƌ��Ȋ����B�����������Ȃ��Ȃ��Ă�̂͑̒��s�ǂȂ낤���B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@�[���܂ł���₱���Ɛ��K���z���Ђ˂���[�ł��Ȃ����[�ł��Ȃ��ƔY�ށB�Y�܂����B
�ŁA��A���˂ɐ��K���z������Ȃ����ƂɋC�������B�ʐς����Œ�ӂ̒�����ϐ��Ƃ���̓ӎO�p�`�������āA�j�b�`�����L���Ȃ�ƍ������Ⴍ�Ȃ�Ƃ��������ʈ���trade-off���L�q�o����Ƃ������Ƃɍl���������B���Ƃ��Ɓi���������ʁ��ʐρj�����ː��K���z�A�Ƃ����ȏ�ɂ͓��ɍ����̖������z�`�Ȃ�ŁA����Ȃ�ʐψ��̎O�p�`���`���ɂ��Ă��������Ƃ��B
�@���K���z�̕ϊ�����Ԗʐς̌v�Z��炢�낢��Ƃ�����ď������̂����A�����؍����g��Ȃ��Ȃ��Ă��܂����B�����B���̂��������Ɏg���@����邩�ȁH �Ȃ������B�Ƃ͂����A����ł��ׂ����Ƃƍl�����͂͂邩�ɃV���v���ɋL�q�ł������A�����Ȃ��B
�@�A�C�f�A�����āA�{���͑��߂Ɏ���ɓP���B
�@���ߑO�A�������B
�@�R�[�f�B���O�����B�[���A�֓lj�B�߂��Ă��đ����B
�@"The specialization v.s.�i�ȉ���)�A�Â��B
�@��Ԗ��ȔɐB�����x�̌v�Z�Ƃ���͌�B�ЂƂ܂��A�a�̎��Ɛۂ��p�x���S�������́i�܂艽���ǂ�����Ă��̂̓K���x�ɍ����o�Ȃ��j�����ŁA�a�̗��p�������ׂē����l�̌�(500)�ł���͂��߂ĂP������܂ʼnĂ݂����ʁi�a�𗘗p�ł��镝�ɂ��Ă̌̐��p�x�̐��ځj�B���͐��K���z�̕��U�̑傫�������ނɕϊ�����Ƃ�����o���ĂȂ��āA���ۂɂ͑S�ē����a�̗��p���̏W�c����u����ށv�̉a�𗘗p���邩�ɂ��āA���p�ł��鐔�ɒ��ɕψق��N���Ă�̂����ǁA���̎��_�ł͂܂������R�g���B
�@������ƕψٗ������߂ɐݒ肵�Ă��Ō��\�K�^�K�^���Ă��āA����������Ƃ��ꂢ�ȃ|�A�\�����z�i�����_���E�H�[�N����Ƃ��̕��z���ă|�A�\���ŗǂ������H�j�ɂȂ邩�Ǝv�������ǁA����Ȃ��̂��ȁB�f�ʐ}�̕������Ă݂�ƃs�[�N���̂��̂��V�t�g���Ă��肷�邵�B�܂��ɓT�^�I�ȃ����_���E�H�[�N�E���f���Ƃ����������B���̂Ƃ���A�a������ސH���Ă������b�g���f�����b�g���Ȃ��̂Ń����_���E�H�[�N�ɂȂ�B�ꐢ�゠����̉a��킠����̐��Ɍ��肪��������Ƃ��Ȃ�generalist���L�܂肻�������A����Ȃ��ǂ��a������悤�ȏ��ł͈�ԗǂ��Ƃ���ɊF�W���������B
�@���̎��_�ł͂܂����ϒl�̕ϓ����l������Ė������A�œK�ȉa����̐��K���z�����l���ĕ��U�̑傫��(���j�b�`���̐���)�������l����������₷�������B���A�ʖڂ��B���ꂾ�Ɓu�����̔䗦�ɋ߂����̂�H�ׂ�悤�ɃV�t�g����͂��H�v�Ƃ����̂��m���߂��Ȃ��̂��B���f���̌������ƍ�Ɨʂ̌y����}�낤�Ƃ��āA��i�̂��߂ɖړI��Y�ꂩ���Ă�ȁB��u�łࢃC�P�邶��Ȃ����v�Ƃ��v��������Ԕ������B���Ă�Ƃ������Ƃɂ��č����̓����B
�@�ŁB�����O��R�̍�}�ɏ�芷����Ƃ��]���Ă����Ȃ���A���܂�gnuplot�ł̏o�͂������肷��B
�@�Q���B���A�������B
�@����ɖ߂��ă��f�������̑���������Ă��璩�ɂȂ��Ă����B
�@�ߑO���A���삳��ɂ�錤���v��B�O���̃��r���[�ɏd�_��u���A�����v�掩�̂͂��ꂩ����܂��Ƃ������ƂŌy���Ƃ����͓_�z���B�Ȃ̂����������ꑫ��тŁA���ɘb��m��Ȃ�B4�̐l�X�ɂ͕t���Ă��̂���ς������̂ł͂Ȃ����Ǝv���B
�@�Z�~�i�[���I����Ă���o�C�N���B�u���[�L�p�b�h�����ׂ����Ƃ̘A�����������̂Ō����̂��߁B�t�����g�͍ς̂����nj��̓u���[�L�L�����p�[�̕Б����K�ƃV�[�����i��łČ�������H�ڂɂȂ����B���i�𒍕����ē������搔�����@�Ƃ̂��ƁB�Ȃ��Ԍ�����ȍ~�̂Ƃ��́A�ЂƂׂ��Ǝ����o�鈫���Ăсc�H�@�A��������������N�̂��̎����ɒ[���������悤�ȁB
�@���߉߂��߂��Ă��ăR�[�f�B���O�c�ׂ̈ɒ��ׂ��̂��Ă�r���Ō����B�ߌ�߂��ċN���B
�@"The specialization v.s. generalization in diet stoichiometry"(����)�A���B
�@GA�̎菇����������Y��Ă���B����܂ł̃��f���ł͑S�������Ŏg���ĂȂ���������w���̏��̎��Ɓi�����̓J���^�搶���S���������j�ŐG���Ĉȗ��c�������H �Ƃ�킯�Ń��f���̏����t���[�������߂��B
�ȏオ�ꐢ��ł���𐢑㐔�����J��Ԃ��B�Ђ˂�̖���trad��GA�Ƃ��������B���������O�̔ɐB�����̌v�Z���߂ǂ��A�c�ƁB
�@���߂���A�������B
�@�������܂Ŏ����ƁB�N���Ă��猤�����B�ߌ�߂��o�C�g�B�߂��Ă��ăR�[�f�B���O�����B
�@��x���A�Óc�N����cgi�̕s��Ɋւ���₢���킹�B
�@�A���̎��䌤�̖q��N����A�������������B�ЂƂ͌����p��cgi�̕\���������������Ƃ������̂ŁA����͕ϐ��̏����ԈႢ�������B�����ЂƂ��v���I�ŁA����cgi�ŕύX��I�����ēo�^���e��ύX���Ă��A�ύX���ꂽ�|�̃��[���͑��M�����̂������ۂɃT�[�o�ɂ��̓��e���������܂�Ȃ�(�ύX�����f����Ȃ�)�Ƃ������́B�������͌������u�ԂɌ��̋C���������B�K���܂��o�^�l�������Ȃ��āA���̂Ƃ���قƂ�lje�������������̂����߂Ă��̋~�������A����Ȓv���I�ȃo�O������Ȃ���ғ����Ă��̂��B��⊾�������Ȃ����}���ŏC���B
�@�������A�e�X�g�ʂ����Ȃ������̂��@���Ɍ����Ă�Ȃ��c�B�܂������B�����ɂł����Z�߂����Ă���}�L�m����̂Ƃ���ɕύX��Ƃ��ǂ̂��炢����������₢���킹�ɍs���āA���ɑ��ɂ��ύX��Ƃ����݂��l��������Ώ����˂B
�@"The specialization v.s. generalization in diet stoichiometry"(����)�A���B
�@�R�[�f�B���O�ɕK�v�ȏ��𗅗ă����߂��B
���Ɋ��̕ϐ��A�c��Ԃ����������ł͂��������Ă��ꂾ�����H
���āA���������B
�@�����N���B���ߑO�A�������B
�@�Z�~�i�[�O�ɏ������Ԃ����������ŁA���̑O����̂�n�߂��i���w��̓o�^��\������X�N���v�g�������Ă݂�B������݂�Ƃ��̂Q�T�Ԃł킸���Q�O�l�サ���V���ɓo�^���Ė������Ƃ��킩��B
�@�ߌ�߂��Z�~�i�[�i�}�j�B
�@�㔼���蕔��B4�̐����N�ɂ�錤���v�攭�\�B�A�H�ҁi�~�W���R�j��C:P��͏]���]���Ă����悤�Ɋ��S�Ɉ��ł���A�Ƃ����킯�ł͂Ȃ��B�ނ���H�ׂ�a�̎��ƗʁA���̑��̗v���ɂ���ĊܗLP�ʂ��ς��͂��Ƃ��������𗧂ĂāA�܂����ɃV���v���ȏ�C/P�ɂ��Ẵ��f���ɂ���Ă��������������Ŋm���߂�A�Ƃ������́B
�@"The specialization v.s. generalization in diet stoichiometry"(����)�A���B
�@���낻�뗈�T�̃Z�~�i�[�Ɍ����ď��������˂Ȃ�Ȃ��B���ʂ��o��̂����肬��Ԃɍ����Ă����荞�݂��Ȃ��Ǝv���Ă����ǁA���Ⴂ���ĂĖl�̒S���͗��T�ł͂Ȃ��čė��T�������B�ʔ����b�ɂ��邽�߂ɂ́A���t�F�C�Y�͌�������������Ȃ����ǁA���Ȃ��Ƃ����t�F�C�Y�̌��ʂ��炢�͏o���Ă��������Ƃ���B
�@�������炭����{��ǂނƉ��̂����ǂ̖{���Q�������Ă���Ƃ����T�C�N���������A�{�i�I�ɐϓǖ{�̍������G���C���ƂɂȂ����B�����������Ƃ������āA����͓ǂ�łȂ����̂̂����t�B�N�V��������W�߂đ������牄�X�Ɠǂ݂ӂ����Ă����B�v�Z������A�N���Ă���Q��܂Ŋۈ���ȏ㉄�X�Ɩ{��ǂނ����̐����𑗂����킯�ŁA���ɍK���Ȉ���������B
�@���ߍ���ł����{�̂����n���J�����̂͂��炩�����炰���B�x���K���A�[�h�V���[�Y�����s���܂őS�������B
�@�����N���B�ߑO���A�������B
�@�i���j���H�����f����"The specialization v.s. generalization in diet stoichiometry"(����)�A���B
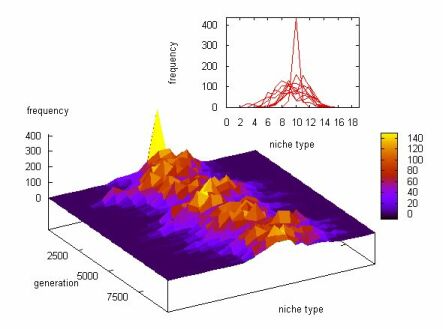
�@�ɐB�����x�v�Z�i�K���x���j�Ɋւ��Ă��ꂱ��B�悭�l������|�������ł̖ʐς���ɂP�ƂȂ鐳�K���z�̊m�����x���Ȃ̂ŁA�킴�킴�y�i���e�B��ʂ��čl���Ȃ��Ă��ǂ��āA���삪�L����ΎR�̍����͏���ɒႭ�Ȃ�̂������B
�@���āA�ŏ��̈Ăł́A�ψقɂ���Č��܂����̂��Ƃ̐��K���z�m(��,��2)�̊m�����x���Ɋ�Â��āA���ꂼ��̉a��(=niche range)���Ƃ̗��p���ifor i=1...n, U(i)=k�P(i<=U<i+1), k�͓K���Ȑ��l�̌W���܂��͊������A���ꂪ�y�i���e�B���x�ɑ�������j��ϕ��v�Z�ŎZ�o���悤�ƍl�����B�����A����́B�ꐢ�ゲ�ƁA��̂̑S�Ẳa�ɂ��Ă̗��p����ϕ��v�Z����K�v������i�����Ƃ����t�ȕ��@�ł���Ԃ̑�`�v�Z��v����j�̂ŁA�v�Z�ʂ��肪�Ƃ�ł��Ȃ��c��ɂȂ肷���Č����I�łȂ��B���̕��@�ł͖����B
�@�ډ��A�c���Ƃ���̉ۑ�Ƃ��ẮA�ŏI�I�Ɍ̂��Ƃ̃j�b�`�����ǂ��W�v���邩�A�A�E�g�v�b�g�̌`�����ǂ�����̂���Ԕ���₷���̂��A�����Ȃ̂����ǁc�B
�@���N���B�ߑO���A���d���B�ߌ�߂��A�������B
�@�i���j���H�����f���A���B
�@���f���̃A�E�g���C���B���f���Ƃ��Ĉ�`�I�A���S���Y���̂悤�ȗL�������̏W�c�T�C�Y�����肷��B�W�c���ŁA�����`�q�^��������łǂꂾ���u������邩���v�Z����B��̓I�ɂ́A�̂��Ƃ̔ɐB���������Ɏ�����ɕ��������̂Ƃ��̕����������܂�B�̂̔ɐB�������x��(1)�H�ׂ��a��stoichiometry��(2)�j�b�`���̍L���ɂ��y�i���e�B�����肷��B�܂��A�̂̕������Ɉ�藦�ň�`�q�ɓˑR�ψق��N����
�@�̂��Ƃ̌`���̍��ق͉a�����̗��p�Ɋւ��镔���̂݁B����͓̌`���Ō��肳�ꂻ�ꂼ��Ɨ��̈�`�q�Ɏx�z����ψقɂ���Đi������B��܂��ȃf�U�C���͍���Q�ƁB
�@(1)�H�ׂ��a��stoichiometry�ɂ��āB
�@(2)�j�b�`���̍L���ɂ��y�i���e�B�ɂ��āB
�@��Ԃ��l��������t�F�[�Y�ł́A�m��Fi=1/n�Ɖ��肵���������A���i�q���f���Ɋg������A����̂��Ƃ̒T���͈͓��ł̑��Δ������Ƃ��Čv�Z�����B
�@���ݍ�p�̕��G�����l������ŏI�t�F�[�Y�ł́A�Œ�̉a���������肵���������A�S�Đ����Ƃ��ċL�q�����悤�Ɋg�������B
�@���N���B�ߑO���A�������B
�@�[���֓lj�(�J)�B�����ނ˖��Ȃ��i�Ǝv���B�I����Ă��炵�炭��������ƎG�k���B��A�������ɖ߂��Ă���R�[�f�B���O�B
�@�i���j���H�����f���A���B
�@�w�ŋ߂��傭���傭�L�������f�B�A�ɍڂ�n�߂܂����B�x�Ƃ������Ɛ��̃L�N�`�N�A�䊈�����̗l�q�łȂɂ��B
�@���A�������B
�@���߉߂�����}���فB
�@�ߌ�߂��A�������ɖ߂�B
�@�[���ȍ~�A�{���͌������̐l�����x�������B
�@�i���j���H�����f���A���B
�@���ߍ��A�������B
�@�l�̌��݂̍�}�X�^�C���́AC�Ōv�Z����gnuplot�ō�}����̂��Aperl�ň�A�̏��������������Ă���Bgnuplot�̂�����R�Ɉڍs�������Ƃ����~�����ȑO���炠���āA�������A�C�}�C�`��芷���铥��肪���Ȃ������B
�@�ŁA�ߌ㒆R�����Ă݂邩�{��ǂނ����Ă݂āAR��}�̈�ۂƂ��ẮAgnuplot�݂����ɑS���R�}�S�}�Ǝw�肵�Ȃ��ł��A�����������K���ɒ������Ă���āi�ꉞgnuplot�����������͂��Ă���邪�j�A�ׂ����w�肪��������Βᐅ���ł��Ƃ������Ƃ��B���̎���ȂǁAgnuplot�Ɋr�ׂĊi�i�Ɏ��R�x�������̂Ƃ��̕~�����Ⴂ�i�����h�N�T�N�Ȃ������j�̂��l����ƁAR�̍�}���͏�芷���邾���̉��l���\���ɂ��肻�������B
�@�܂����K�f�[�^���g���Ă݂Ďd�l�͈��߂��i�悤�ȋC�ɂȂꂽ�j��ŁA���͎��ۂɎ����̖��ɗ��ĂĎg����������Ă݂�Ƃ��납�ȁB��肽�����ƂɎg���Ă݂�A���I�ɏ��P���}���Ă݂���A���̋t�ɏo���Ȃ����Ƃɕs�����o�Ă����蓙���邾�낤�āB�����Ƃ��A���߂œ��v���g���p�r�����������܂ō�}�����Ȃ�ŁA��Ԃ��������Ƃ��낪�g���ĂȂ��̂����B
�@���N���B���ߍ��A�������B
�@�ߌ�߂��A�Z�~�i�[�i�}�j�B�O���͐蕔��B4�̐ΐ�N�ɂ�錤���v��B��̕t�����ނƑ��ސH�ҍ����̑��݂̉e�������悤�Ǝv���܂��A�Ƃ������e�B�܂��܂���͊|���肻������������_�����B���\��ɁA�Ȃ���Ȃ̂��Ȃ����������ł��Ȃ��̂��Ƃ����w�E������Ă����B���߂��Ɏ��O�������Ă݂��Ƃ����t�B�[���h�i�t�̐X�j���A���̗v�����������ĉ������Ă�̂�������Ȃ��Ȃ肻�����Ɗ����������ɁA����͂Ȃ��Ȃ��ǂ����������B
�@�㔼�͐�t��B4�̖ؑ��N�ɂ��_���Љ�B�������̂����A�����Ȋ��z�������Ώ��X�s���ȓ��e�������B�͓c���ɂ�����ߏ�Ƃ�������Z�~�i�[�O�̃P�A�́A�����ɂ킽�鎎�s����̖��ɂ���`�Ԃ����A����ɂ���ăZ�~�i�[���ɍő���̋c�_���o����悤�A���O�Ƀv���[���e�[�V�����̎����ψ�ɒ�グ����w�͂��}���Ă���B���̂���B4�̔��\���Ƒ�T�����i�K�łقƂ�Ǘ��ꂪ�������Ă��āA�����͏\�S�ɐi�ނ��c�_���̂��̂�����ފ����͂�┖���i�悤�ȋC������j�B
�@���N���B
�@�I����Ă���o�C�N���B�O��u���[�L�p�b�h�ƃG���W���I�C�������̌��ς���B�p�b�h�������i���A���\�H���������āA�S�����݂łQ���~��B���\�������������n�͂���ȏ�扄���Ƃ�����ɂ������Ȃ����A�����ł��̂��|���B���ʂ̂ŁB�I�C���ʂ͎��O�Ō�����Ƃ��Ă������̂����A�������̍H���ݒ�͈�����ł��łɂ���Ė���Ă��l�i�I�ɕς��Ȃ��B��Ǝ��Ԃ͂��قNJ|����Ȃ��Ƃ������Ƃ��������ǁA���T���Ɏ��Ă�����āA�y�j���Ɍ�����ƂƂ������ƂŎ�z����B
�@�ߌ�߂��A�������ɖ߂�ƉY�ˎ��K�ɂ����Ă��l�X���߂��Ă��Ă����B������D�V�Ƃ����킯�ɂ͂����Ȃ������悤������ǁA����ꂳ��ł����B
�@�_�����e�����FCNR�ɂ�鋣���r���x�����f���A�Â��B
�@���N���B�ߑO���A�������ƁB�ߌ�A�������B
�@�悲��̗�[�̌��A���ׂ���Ό������̎��������g�[�������Ƃ����I�`���B�����@�����ʂȂ̂ŗ������̕������g�[�ɂȂ��Ă���Ɨ�[������Ȃ��Ƃ������������������d�l�Ȃ̂����A�������Ƃ����ʂƂ͎v��ȂB���̎����ł����Ƃ����ԂɂR�O�x�㔼�ɒB����̂͂܂����B�Ăǂ����悤�B
�@�I����Ă���E���x����ɌĂ�āA�ǂ�Œ����Ă�_���Ɋւ��Ă��ꂱ��Ǝ������B���Ɋ�����P�ĂƐ����s���ŏ����������K�v�ȉӏ��̎w�E��Ⴄ�B���炭������������A�g�K�V�N�̏C�_�����ɂ��Ă̘b�������肵�Ė�O�Ɏ����B�������ɖ߂��Ă��đ������ꂱ���ƁB
�@gnuplot��timefmt���ǂݍ��݂Əo�͂łȂ����ꃖ�����������ہB�T���Ƃ��ēǂݍ��ނƂU���Əo�͂����A���������B
�@�i���j���H�����f��
�@��Ԃ������Ƃ��̘b�͂��قǎ��Ԃ��|�����Ɍ��ʂ��������������B�����͂��̃Z���Ői�߂Ă������B
�@���N���B�ߑO���A�������B
�@�i���j���H�����f��
�@���N���B
�@��������C���I�Ƀ��t���b�V�����Čߌ�x���Ɍ������B�����̒������BMcCann et. al.(2005), "The dynamics of spatially coupled food webs", Ecol.letters(8):512-523.���I���V���������B
�@�i���j���H�����f��
�@�����F�hgoogle�d�샊�t�@�����X�h
�@double�^�̊ۂߌ덷�̂��Ƃ����O���Ă��đ啪���Ԃ����X���Ă��܂����B���_���炢����stl::map
�@���ǁA���炩����int�^�Ɋۂ߂�stl::map<>����߂āAdouble arr[int]�ő�p�B����ł悢�Ƃ����̂ɋC�Â��܂Ŏ��Ԋ|���肷���B
IBM�ŋL�q�����u����Ҍ̂����ׂ��p�����[�^�v�́A
�܂��萔�A
�˃j�b�`���̕��ϒl�i�̐������Ɉ�`�q�^����Z�o�j
�˃j�b�`���̕��U�i�̐������Ɉ�`�q�^����Z�o�j
06��13��(Mon)
�@�{��ǂ�ł��猤�����B�ߌ�߂��܂ŃR�[�f�B���O�A�ߌ�߂��Z�~�i�[�i�}�j�A���ǂ��Ă��ăR�[�f�B���O�B
�@�X�N���v�g�̒��g�͂����������e�ł͂Ȃ��A���v���Ԃ͂P�O�����炢�������̂����A�P�O���O����n�߂���Ŋ낤���Z�~�i�[�ɒx������Ƃ��낾�����Bgnuplot��timefmt=date�łȂ������̕\�����ꃖ�����Z����Ă��܂����͉������Ė����āA�ꂵ����ɓ��t�����o���Ȃ��ʖڎd�l�B
�O������t��B4�̐��N�ɂ��_���Љ�B�������̂����A�S�N���̍��̖l�̃v���[���e�[�V�������霂Ƃ�����A����߂ăA���Q�ȓ��e�ŃZ�~�i�[�Ƃ��Đ������Ȃ������B���̂��܂�̓��e�ɐ�t�����{���āA�ė��T���炢�ɍēx��蒼���^�тƂȂ����B������Ɗw���ׂ̈��l���Ă�؋��ł͂�����A����ȃR���C�̂͏��߂Č����B�c�Ƃ䂩�A�u�l������Ȃ������Ȃ��v�Ƌ���������Ă��̂ŁA�������{���Ă�悤�ŋ�������Ȃ������̂������B
�@���f�����̂͊ȒP�ɂ����Ƒ�Ӄ��X�̕ω��ɒ��ڂ������̂ŁA�ڍׂ͏Ȃ����A�C�f�A�Ƃ��Ă�NC/P=(AC-R)/AP�Ƃ��������B���ɃV���v���ł��邪�A��ӗ�(R)�ɂ��Ď������Ă����Ɖa�̗ʂƎ��̗��ʂ������҂̑̓��ܗLC/P������ł��A�c�_�ł�����e�͋ɂ߂đ����B�����̕��͉a�̎��E�a�̗ʁi�{�����̂������j�ɒ��ڂ���Ƃ̂��ƂŌ��ʂ��y���݂ŁA���ɖʔ������e�������B
�@�Z�~�i�[���I����Ă���A����s���ȓ_�𐴐��N�Ɏ��₵�����A�ǂ������Ă��āA������ɂ����Ȃ�I�m�ɓ����Ă��ꂽ�B
�@�A�E�g�v�b�g�͊�{�I�ɔ��q�Q�}���g���ĕp�x���z��������悢�A�̂��ȁB�{���͂Ђ�����R�[�f�B���O�B�̂̃j�b�`���̕����ɂ��āB���[�Ƃ��I���Ȃ��B
�@�C���g���_�N�V�����̍\���̕��͂��낻��l���n�߂āA��������̂܂ܔ��\�̍\���Ɏg���Ă��܂����B
06��12��(Sun)
�@�����̓��j���p�^�[���B�ߌ�߂��A�u���[�L�p�b�h�̌����Ƀo�C�N��ɕ������A�����ׂƂ̂��ƂŐ摗��B�����B
�@�w(��)�`�x������ɍ���Ȃ��Ȃ��Ă�����łƂ肠��������B�ŏ���"Do you wanna be the eater of ONLY-ONE/ANYONE ?"�Ƃ����̂��l�������ǁA������ƃZ���X�����ăJ�b�R�����̂Ŗv�B������ǂ��ɂ������������B
�@���āA����ł͂ǂ����邩�H�@�����͎��ɃV���v���Ōv�Z���Ȃ���Ηǂ��B���Ȃ킿�A�m���ϐ�U��W�����K���z �m(0,1) �ɂ��������m���ϐ� Z=(U-��)/�� �ɕϊ����Ă���W�����K���z�\���Q�Ƃ���悢�B����Ȃ��Ԃ̍����v�Z�����ōςށB�W�����K���z�\�͂��炩���߃e�L�X�g�f�[�^�ŗp�ӂ��Ă����āA�v�Z�J�n����map(double z -> double val)�Ƃ����`���̃n�b�V���e�[�u���ł�����Ă����ΐϕ��v�Z�����̃I�[�o�[�w�b�h��S�ĉ����ł���B����Ȃ猻���I�Ȍv�Z�ʂŎ����ł��������B
06��10��(Fri)
�@���d�����Ђ��Ă���o�C�N��Ɋ�蓹�B���Ă��u���[�L�p�b�h�������Ă邩�Ǝv��������גx��Ƃ̂��Ƃœ��j�ɂ��ꍞ�݂����Ƃ̂��ƁB�����Ȃ��ŋA��̂��Ȃ��A���������Ȃ�ŃG���W���I�C��������Ɍ����B����͍���]�p�̃Z�b�e�B���O�B��߂̃M�A�̂܂�܂ň��������Ă��G���W����������Ȃ��̂ł��炭�C�������ǂ��B
�@���f���̑g���Ă̎��s����̑����B�A�C�f�A��F�X�ϑz����̂ɑ����āA���̒i�K�����Ɋy�����B��܂��Ƀf�U�C�����ł����̂ŁA���Ƃ͒���������B
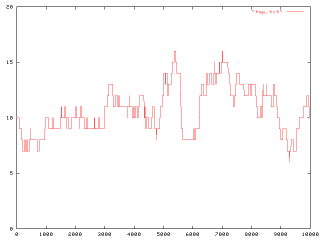
�@����ԃI�[�o�[���b�v�͍l�������A������̌̐����W�c�T�C�Y�����ꍇ�ɂ͏W�c�T�C�Y�Ɠ������Ȃ�܂Ń����_���Ɍ̂��Ԉ����B�t�ɏW�c�T�C�Y�ɖ����Ȃ������ꍇ�́A�c�ǂ����悤���ȁA�����������ɂ�����������W�c���̂��̂����ނ���Ƃ��Ă������̂��ȁB�܂��A�����͗^����p�����[�^���悩�B
�@�ŏI�I�ȃA�E�g�v�b�g�Ƃ��āA�����Ԍ�ɏW�c�̍\�����ǂ̂悤�ɂȂ��Ă��邩���ώ@����B
���iQ=R/C�j��Q1<Q2<�c<Qn�ƂȂ�悤�Ȏ����Q���l���A���ꂼ��̉a�̑��Δ������i�����₷���j��F1,F2,�cFn�Ƃ��ė^����B
�@����̂��ǂ̉a���ǂꂾ�����p�ł��邩�����g�̊m�����x���z�ŋL�q�����̂̃j�b�`���Ɋ�Â��Č��肷��B�������A���̕��z�`�͐��K���z�`�����肵�A���ςƕ��U�����ꂼ��Ɨ��̌`���l�ƍl���āA�Q�Η���`�q�E�����ʁE�����̂̈�`�q�Ɏx�z�����B����ai�̗��p����Ui�Ƃ���B�܂�Ui=0�͔��������ai�����̌̂ɂ͐H���Ȃ����̂��������Ƃ�\���B
�@���A����҂������ɗv������h�{������ƂƂ���ƁA�ai�ɂ��Ă̓]������Qi/�Ƃ��l���邱�Ƃ��o���A�ai����l�������Ƃ��̗��v�͔������Ɨ��p���Ɠ]�������̐ςɂȂ�B���Ȃ킿�A�oF�U�Q/�Ɓpi�B
�@�S�Ẳa�̔��������������Ƃ���ƁA�a���Ƃ����Ƃ��ɉai��������m��Fi=1/n�B���ɂƂ�a�������_���ł���Ɖ��肷��ƈꐢ�゠����ɉa��N�Ƃ����Ƃ��̗��v�́A��(i=1...N)�oF�U�Q/�Ɓprand[1,n]�ƂȂ�B
�@�a���p�Ɋւ��閧�x���͖ʐς�1�Ƃ��邽�߁A���U���傫���Ȃ��ė��p�ł���a�����̕����L���Ȃ�ƁA���̕��A�a������̗��p�����͒ቺ����B
06��09��(Thu)
�@�{��ǂ�ł��猤�����B���ǂ̖{���������܂��Ă��邹���������Ėڈڂ肵�Ă��܂��B���ǁA�������̖{�s���ēǂ�ł���̂����ǁA���g�̍D�݂��猾���Έ������O���ēǂ܂Ȃ��ƎU���ȋC�����Ă��܂��B�����͂����A�S���ʂ̕���ɂ�������炸����ɋ��ʂ������O���ł����肷�邱�Ƃ������āA�ŋ߂͂��������̂��ʔ����B
�@�܂��͋�ԂȂ��̏����ł̃V���v���i�ɂ������j���f���̍\�z�����ꂱ��B���K���z�̌`�����肵���`�����ǂ�����Đi�������悤���Ƃ��ꂱ��Y��ŁA���傤�Ǘ֓lj�������Ƃ������āA��������t���ɑ��k���Ă݂�B���낢��Ƌ�̓I�ɂǂ��������邩�Ɋւ��ăA�h�o�C�X�����������āA����������B
�@���ǁA���K���z�̕��ςƕ��U��Ɨ��̌`���Ƃ��đ����āA���ꂼ��ɑ����ʂɂ�鑊����`���U�����肵�ĂƂ肠�����������Ƃɂ��āA�܂��̓E�H�[���A�b�v���Ă�ߋ������Ă݂悤�B
�@����������Q�Η���`�q�E20���ʁE�����́i1/0 * 20 * 1)�ɂ���āu���K�������肳�ꂽ����`���v�̕��U���x�z�����C�L���m��10^-4���x�̕ψّ��x�����Ƃ��ɁA���S�ȃ����_���h���t�g���������肷��ƏW�c���ɂǂ̂悤�Ȍ`�����L�����Ă�����������Ă݂�B�ŁA�ˑR�ψقɂ���āA���Ԍo�߂ƂƂ��Ƀj�b�`�����L�������苷�܂����肷��e�X�g�Bok�A���������̐v���o�͒͂߂��B
06��08��(Wed)
�@�ߑO���A�Z�~�i�[�i�J�j�B�{���̒S���̓}�c�V�}����ŁAD�_�̍\���ɂ��Ă̔��\�B�����ނ˂���܂łɒ����Ă����b�B�Ƃ͂����A����܂ł̏������悻�S�Ē��锭�\���e�������̂Œ����̂��^�C�w���������B
�@�s�v�c(�ɕ�������)���肪�������o�ꂷ��̂��C�ɂȂ����B���������ꂪ�Ó��Ȃ̂��ǂ����A�l�ɂ͂�����Ɣ��f�ł��Ȃ������̂����ǂ������B�������Ƌ��d��justification�����Ă����c�b�R�~���ł����̂�������Ȃ��B���A�������A��O�̃f�[�^�T���v�����O�ɂ��ĕW���Ŗ������肪�g���Ă����Ƃ��ɁA���̑Ó����̗ǂ�������������ƕ]���ł��Ă��Ȃ��i���̎�@�ɐ��荞�܂ꂽ���肪�����Ȃ�ǂ����ǂ����������w�E�ł��Ȃ��j�̂ŁA����͒�����Ƃ��Ă̖l�̕��ɂ���肪����B
�@��������������Ă����ēǂށBWilliams and Martinez(2000), Nature(404):180-183.��Kawata(2002), Proc.R.Soc.Lond.B.(269):55-63.��ǂނ��A�O�҂�simple��搂������\����c�B
�@�߂��Ă����Ƃ���ŁA�R�{�N�ɑ��܂�B�Ȃ�ł�������l�b�g���[�N�ڑ����s���炵���B�F�X�ƒ��ׂĂ݂Ă������I�ɂ��_���I�ɂ����Ă���`�Ղ������A�ǂ���琶���n��DHCP�T�[�o�Ɍq����Ȃ��H�A�Ƃ������_�ɗ��������B�������A������ۂ�����]���ŕ����Ȃ��̂ʼn����T�[�o���̖��Ƃ��Ⴄ�悤�ȋC�����āA�ǂ����������C�}�C�`����ʁB���ǁA�Œ�IP��U���ĉ�������Ƃ������d��A����ɔ����e��A�b�v�f�[�g���Ȃɂ��B
�@�����͋A���l�X�������������߂ł���B���̂ɐl�X������قǂɃT�b�J�[�ϐ�ւƋ�藧�Ă���̂��ɂ͉���畡���I�v��������悤���B�Ƃ��낪�������ꂽ���l�����ɂ����₤�ƁA�ӊO�Ȃ��ƂɁw�ʔ�������ʔ����̂ł���x�Ƃ������������Ԃ��Ă��Ȃ����Ƃ������B�i�������ؓ����ĂČ��l������ɂ͋��邪�j
�@���Ƃقǎz�l�ɐl�X���䂫���Ă�܂Ȃ����̋��Z�̖��͂����邱�ƂȂ���A���̋��Z�ɂ������䂩��Ă�܂Ȃ��l�X���i�����炭�͂��Ȃ蕁�ՓI�Ɂj���݂��闝�R�ɂ��Ă��ꂱ��l���Ă݂�̂��l�I�ɂ͖ʔ����B
�@���āA�ǂ������G�������b������Ă������A��܂��ȕ������͏o���Ă����B�ŏ��͋ɗ̓V���v���ȃ��f�����g���Ă���܂ł̗��_�̂����炢�Ɠ��ݑ�����B���̃X�e�b�v�Ƃ���Kawata(2002)�Ɏ����a�̒T����Ԃ��l�����郂�f�������B�ŏI�t�F�[�Y�ł�Williams and Martinez(2000)�̂悤�ȐH���ԃp�^�[���`���i���G���j�ɂ��Ă݂Ă����B�c������Ɨǂ��Ȃ��B
�@(��)�c�Ƃ����A�ŏI�I�ȗ��Ƃ�������������c�_�ł������Ȃ̂͂��ꂵ�������B
06��07��(Tue)
�@�v�X�ɖ�X�������Ē��܂Ŗ{��ǂ�ł�����A���̂܂ܐQ�߂������B�ߌ�߂��o�C�g�B
�@�|������̃T�C�g�ɂāAR �Ńv���O���~���O�F�f�[�^�̈ꊇ�����ƃO���t�`�������J����n�߂��B������@��R�Ɉڍs���Ă݂悤���Ƃ������ƂŁA����������@������Ă݂�B
06��06��(Mon)
�@�ߑO���A����œǏ��B���[�ށA�}���ق܂œǂ݂ɍs���ق����i�����ǂ��ȁB
�@���߉߂�����A�_�����������B�Z�~�i�[���͂���Ŗ�܂ŁB�w�E�̂������ӏ��ɂ��đS�đΏ����Ă��烁�[���ōđ��B��A�܂��͋�ԂȂ��̃��f������f�U�C���J�n�B�v�������V���v���ȍ\���ɏo���邩���B
�@�ΐ�N�͂��܂��������Ă��Ȃ��������ǁA����Ƃ͕ʂɖl�̌����e�[�}�ł���A��ԂƐ��̓������l�������ށ|���ސH�Ҋԑ��ݍ�p���l���邤���ł��A��O�ɏo��̂Ƃ͕ʂɁA����i���̓����j�݂̂��o���邾���V���v���Ɋώ@���邽�߂Ɏ��������n��g�ނ̂̓A�����ȂƊ������B
�@����Ƃ͑ΏƓI�Ƀ}�N���̃Z�~�i�[�ł́A�قƂ�ǂ����������P�A�͖����A���̏�̔��\�҂��Ƃ̃v���[���̓����ʂƔ\�͂����̂܂ܔ��f�����B���̑���A�����Ă鑤�Ƃ��Ă͊w��\�Ɠ��������̏�̏������ɗ������Ď��^��������Տꊴ������B
�@�ǂ��炪�ǂ����Ƃ�����ł͂Ȃ��A�꒷��Z�Ƃ��������Ȃ����A����͂�����ƃ��X�N���ɌX�������������������B
06��05��(Sun)
�@�I�t�B�V�����T�C�g���I�s�����ɂȂ��Ă��q�r�L����܂ŊςĂ��猤�����B�����̓��j���p�^�[���B�ׁX�ƍ�Ƃ��Ă��琳�߉߂����p�ʼnw�O�A�ߌ�߂��܂ŁB
�@����ɂ��Ă��A�~�J�O�Ȃ̂ɑO��Ƃ��������āA�A�^���̎��ĂȂ���ԂƂ����̂͏������Ȋ����ł͂���B���A�͌��\�O����w�^���Ă�����A���߂Ɍ������Ă����Ηǂ������B
�@�w�E�̂������}�\�̃��C�A�E�g�ύX�ƏC���A�����s���̉ӏ��ɂ��ĉp�앶�B��܂ŁB
06��03��(Fri)
�@�ߌ�߂��A�֓lj�i��j�B�{����Lampert and Sommer�́wLimnoecology�x�͓Ǘ��B���Ԃ���⒴�߂������A�ꏊ��蕔���Ɉڂ��čŌ�܂ŁB
�@����U�������_���Љ���������ł��Ƃ����H���ύX�B��������͍̂Œ���}�\��abstruct�����Łw���҂ɑ����āx�v���[��30��/�c�_30���Ƃ������ƂŁA���\��vs������̓��_���K���_���炵���B�{���̕⑫��ok���������������Ƃ��ăp���[�|�C���g�Ɠǂݕ��I���W���s�B������Ƃ͎��ԓI�ɂ͂����������Ƃ���������ɁA���A���^�C���ȃv���[���Ƌc�_�̔\�͂��v�������킯�ŁA�Ȃ��Ȃ��v�����x���̍������ȓ��e�����A���Ă��܂���邩�H�@7������ɒS��������Ă������Ȃ��A�_���I�тƂ����O�̔��\�\���̌����Ȃ��C�������Ȃ��Ƒ�S���ɂȂ�\���B
�@���˂ɁA�悳���ȃA�C�f�A���v�������B�J���^�搶�̂�����ƑO�̊i�q���f���Ǝ����悤�ȉa���������݂��镪�z�ƕߐH�҂̉a�T���͈͂��l�������f�����g���āAspecialist v.s. generalist�Șb���o���������B
�@�x�[�X�R���Z�v�g�Ƃ��Ắw�����͎����̃Ƃɋ߂����̂�������A�̂ł͂Ȃ����낤���H�x�B�Ȃ��Ȃ�A���̕����]����������������ł���B
�@����Ƃ��Ă����̂́Ageneralist�i�����̉a�ɑΏ����邽�߂̃j�b�`���̍L���ɃR�X�g�y�i���e�B��^����jv.s. specialist�i�Ώ��튯�ւ̓����R�X�g������������ɐH����a�̕����������j�B
�@�\�f�Ƃ��ẮA��Ԃ������i�������͉a���Ƃ̋Ǐ��I�ȓ�����Ғl���ψ�ł���j�ł͉a�̗��p���Ɋւ���j�b�`�����i=specialisation�ɂ��������j��������B����ɑ��āA��ԓI�ɕs�ψ�ł���ł͂��generalist�I�Ȍ`�������n�������̌n���߂�B�i�����m��Ȃ��j�B
06��02��(Thu)
�@����̕����̕��ʂ̂��ߐt�R���فB������̕����������ČߑO�������ς��Ă����Ă����̂������Ȃ�W���ł����B�c�}���ٗǂ������B�ߌォ�猤�����ɖ߂��Ă��āA����ς�{��ǂ�ł����B���傤�͏I������Ȋ����B
�@�[���A�֓lj�i�J�j�B�S���͂܂��܂��n�ӌN�B�ǂ��ł��ǂ��̂����A�ނ�ln(logarithm natural)���o�Ă��邽�тɁu���v�u���v�c�A�Ɠǂނ̂��C�ɂȂ��ċC�ɂȂ��Ă���[���Ȃ������B�l���g�́u�����������v�Ƃ��u�낮���[�v�Ƃ��ǂ�ł邯�ǁA���Ⴀ�����ǂނ̂����������Ƃ�����ƁA���āH �܂����̂Ă̋c�_�͊�{�I�ɕs�тŁw���x�ł��w���x�ł������낤���ǂ��A�C���I�ɂ͐���肪�����悤�ȁB�u���邦�ʁv�Ƃ�����ƃV���{���b�N�����N�\��̂��Ƃ����C���ɂȂ邵�B
�@�e�핶�������낻�됮�����A�����邱�Ƃƌ����Ȃ����ƁA�o���邱�ƂƏo���Ȃ����Ƃ̃s�b�N�A�b�v�B�iomnivory���܂ށjintraguild predation�Ɋւ���g�s�b�N������قǐ���̍L���e�[�}�Ƃ͓����͎v�������Ȃ������BPolis and Holt(1992)���q�ׂĂ���悤�ɁA�H���H����Ƌ������Ɏ���ɓ���镡���I�Ȑ����ԑ��ݍ�p�Ȃ̂ŁA���ꂾ���ʔ����e�[�}�Ƃ������ƂȂ낤�B�����j�\���ɂ��a�T�C�Y�V�t�g�Ȃ��l����ƃv�����N�g�������̂ւ�ŏ\���Ƀ��f���Ƃ��Ď����Ă��������Ȋ����B
06��01��(Wed)
�@����Ŗ{��ǂ�ł���c�B�{���̓Z�~�i�[���}篎��~�߁A�����A�~�J�O�A12�����_���O�B���ꂼ�V�̔z�܁A����ɂ������B
�@�R�`�����암�ߕӂ����낤��A�ǂ����Ȃ炠�������E���E�����Ȃ��Ŏ����h�̕��֍s���悩���������B���ꂱ��Z�����đ���ɍs�����̂����͍��N���߂ĂƂ������ƂɋC�������B���\�̗͗����Ă邵�B�Ƃ͂����A��͂�T�X���Ă��������肾�Ƃ�낵���Ȃ��B
�@���X�ɃA�C�f�A�Ƃ�������ԑ傫���X�g�[���[�̕������͔��R�Ȃ��猩���Ă����A���ȁB��̓I�ɂǂ�������ǂ��]����̂����C�}�C�`�Z�܂�Ȃ��̂ŁA�܂��܂��v�����܂܂̃A�C�f�A�𗎏������B
�@Polis and Holt(1992), "Intraguild predation: The dynamics of complex trophic interactions", Tr.E.E.(7)5: 151-154 ���t�R�ɂ���̂��B���ʂɍs�����B���낻�땶�����A�^�}�̒��ŖO�a���Ă����悤�ȁB
�@�֗��Ȃ��ǂ킴�킴�ɂ͎g��Ȃ�google�̖��ʋ@�\�̃��t�@�����X�B�ǂ�����Ē��ׂ��̂�����ԋC�ɂȂ邯�ǁA�ЂƂ������Ă������̂��Ȃ��B
{kind=link}
{kind=link}